
Simulium (Eusimulium) guimari Becker, 1908
|
publication ID |
https://doi.org/10.1080/0022293032000140958 |
|
persistent identifier |
https://treatment.plazi.org/id/039787B9-FFB6-575F-69E9-FCDA4A453B17 |
|
treatment provided by |
Carolina |
|
scientific name |
Simulium (Eusimulium) guimari Becker |
| status |
|
Simulium (Eusimulium) guimari Becker View in CoL
Material from breeding sites
Gomera: Site G2—1„(z), 1 larva ( 6 June 1990) ( RWC) . Site G3—9„(z), 6”(z), 151 pupae, 313 larvae ( 6–7 June 1990) ( RWC) . Site G5— 1 pupa (advanced male), 6 June 1990 ( RWC) [possibly guimari / velutinum hybrid, see ‘Remarks’]. Gran Canaria : Site GC 6— 5 pupae, 16 larvae ( 24 April 1996) ( RWC) . Site GC 8—larvae present ( 28 March 1994) ( BM / ANN). Site GC9— 2 larvae ( 28 March 1994) ( BM / ANN) , 2„(z), 2”(z), 4 pupae, 32 larvae ( 30 March 1995) ( RWC) , 2„(z), 1”(z) 19 pupae, 62 larvae ( 1 April 1995) ( RWC) , 39 immatures in sample ( 19 November 1995) ( BM / ANN), 2„(z), 23 pupae, 27 larvae ( 21 April 1996) ( RWC) and 2„(z), 2”(z), 3 pupae, 26 larvae ( 20 March 1997) ( RWC) . Site GC11— 1 larva ( 21 April 1996) ( RWC) . Site GC 12—larvae present ( 28 March 1994) ( BM / ANN). Site GC13—1„(z), 2”(1z), 1 pupa ( 31 December 1987) ( MB) and 3 larvae in sample ( 28 March 1994) ( BM / ANN). Site GC15—3„(z), 8”(7z), 1 pupa ( 26 December 1987) ( MB) , 6„(z), 13”(z), 29 pupae, 174 larvae ( 11 August 1988) ( MB) , 5 pupae, 15 larvae ( 27 March 1994) ( BM / ANN) and 91 and 103 immatures in respective samples ( 27 March 1994 and 16 November 1995) ( BM / ANN). Site GC20— 1 pupa ( 31 March 1994) ( BM / ANN) . La Palma: Site LP2— 1 pupa ( 16 April 1981) ( RWC) [see ‘ Remarks’]. Tenerife: Site T1— 24 larvae ( 12 April 1991) ( BM / ANN / MB) . Site T8— 31 larvae ( 1 November 1991) ( BM / ANN / MB) . Site T9—2„(z), 21 pupae, 26 larvae [these originally misreported as ‘ tenerificum ’] ( 8 April 1983) ( RWC), 4 pupae, 81 larvae ( 14 April 1991) and 4 larvae ( 1 November 1991) ( BM / ANN / MB) . Site T10— 7 pupae, 10 larvae ( 9 November 1991) ( MB / ANN / MB) . Site T14—1 „(z), 1 ”(z), 1 pupa ( 24 March 1983) ( MB) , 1„(z), 16 pupae, 115 larvae [these originally misreported as ‘ tenerificum ’] ( 9 April 1983) ( RWC), 2”(z), 5 pupae, 7 larvae [four infected with mermithid parasites] ( 13 June 1990) ( RWC) and 5 pupae, 99 larvae ( 11 April 1991) ( BM / ANN / MB) ; 12 larvae ( 14 April 2001) ( MB) . Site T23— 45 pupae, 55 larvae, with one larva labelled ‘wet wall’ ( 17 April 1991) ( BM / ANN / MB). Site T24— 48 pupae, 11 larvae ( 15 April 1991) ( BM / ANN / MB) . Site T27— 1 larva
( 13 April 1991) ( BM / ANN / MB). Site T 28— 4 larvae ( 6 November 1991) ( BM / ANN /
MB) and 2 pupae, 147 larvae ( 16 April 1991) ( BM / ANN / MB) .
Remarks
This species, endemic in the Canaries and occurring only in the four islands listed above, is morphologically the most distinctive species of the aureum group ( Eusimulium ). The male is not dissimilar to that of other aureum group species and has very similar genitalia (parts illustrated in Crosskey, 1965), but the female differs by having the tarsal claw tooth reduced to a small peg (Crosskey, 1988b: figure 16), the larva lacks ventral papillae and has a reduced arcuate postgenal cleft (figure 42), the pupal gill filaments are unusually short and widely spread (figure 19), and the pupal head plate and thoracic dorsum are very closely covered with minute microtubercles. Also unusual in the pupa is the form and weave of the cocoon: the anterolateral corners have a cut-away shape (sometimes less markedly in pupae from Gran Canaria than from Tenerife) (figure 27) and the cocoon is loosely perforated near the front margin (figure 26). The single pupa available from La Palma (data above) is even more remarkable for its cocoon shape: the cocoon lacks the usual anterolateral parts and fits the pupa as an unflared sleeve which, seen in profile, has a straight and strongly sloping anterior margin (figure 28). The pupa is immature and the gill filaments, probably on this account, are not widely spread basally (figure 21) or fully straightened out—as the result of which the gill rather resembles that of S. tenerificum . Identification of this pupa as S. guimari is confirmed, however, by absence of any dorsal spine-comb on segment 6 and by dense and even covering of exceptionally small microtubercles on the head plate and thoracic dorsum.
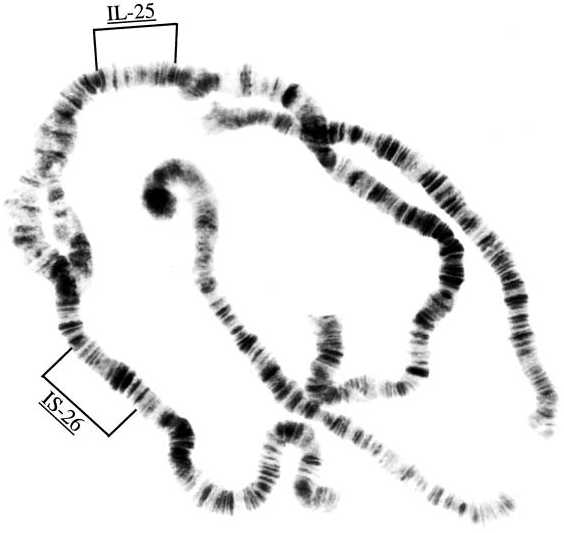
Simulium guimari is the Canarian endemic chromosomal aureum sibling ‘M’ defined and placed in a cytophylogeny for the Simulium aureum group by Leonhardt (1985); it was wrongly referred to by Crosskey (1988b), because of misassociation of the larva with other life stages, as Leonhardt’s sibling ‘L’ (~ tenerificum ). The characterization was based on larvae from the streams in the Barranco del Rio and Barranco del Infierno (Tenerife sites T 9 and T 14) collected ( RWC) on 9 April 1983. These larvae differed from sibling ‘J’, which seems to equate morphologically with S. petricolum Rivosecchi (a species mainly of southern Europe and Madeira) by two fixed inversions, IL-25 and IS-26. Further cytological data for S. guimari were obtained in 1995 when polytene chromosome preparations were made (P. H. Adler) of 14 larvae from Gran Canaria island (data: Barranco de la Mina near Las Lagunetas, 30 March and 1 April 1995, RWC). Figure 44 View FIG shows the polytene chromosome complement with positions of the fixed inversions IL-25 and IS-26 marked. Chromosomes from this Gran Canaria population correspond in every respect to aureum ‘M’ from Tenerife (Adler, personal communication); specifically, the complement differs from that of aureum ‘C’ by four fixed inversions (IS-10, IS-11, IS-26, IL-25, see Leonhardt, 1985: 2059), in male larvae the expanded region of chromosome I is unpaired and no floating inversions were seen.
The fact that populations are chromosomally identical in Tenerife and Gran Canaria verifies cytologically the synonymy of Simulium submorsitans with S. guimari formally established in an earlier work (Crosskey, 1988a: 448). Séguy (1921) described submorsitans from a unique male collected near Tafira in Gran Canaria and illustrated its genitalia. However, both specimen ( holotype) and slide preparation are lost, as discussed in Crosskey (1988b: 341). Nevertheless, the combined evidence derived from Séguy’s drawing, the occurrence of chromosomally identical populations for S. guimari in the two islands relevant to synonymy, and the fact that S. tenerificum (unlike S. guimari ) has never been found in Gran Canaria, clinches the synonymy.
Attention should be drawn to the male pupa listed above from the Gomera aqueduct site G5. This was found sympatrically here with S. velutinum and has features suggesting that it is possibly a guimari / velutinum hybrid specimen. Its characters typical of guimari are: cocoon somewhat sleeve-like (sides not flared) with anterolateral corners cut back, cocoon anterior margin broadly prominent, head plate and thorax very densely covered with extremely small microtubercles, gill filaments widely spread and uppermost filament evenly curved, male genital style shape; its characters typical of velutinum (and also tenerificum ) are: pupal abdomen dorsally with partially developed spine-comb on segment 6, male genital ventral plate in plane view with distinctly flared subtriangular shape (not narrowly blade-like as is typical for guimari ).
| BM |
Bristol Museum |
| MB |
Universidade de Lisboa, Museu Bocage |
| T |
Tavera, Department of Geology and Geophysics |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |