
Buthacus yotvatensis Levy et al., 1973
|
publication ID |
https://doi.org/ 10.1206/0003-0090.450.1.1 |
|
persistent identifier |
https://treatment.plazi.org/id/0396327D-FFF5-7E67-B017-6D2632056B96 |
|
treatment provided by |
Felipe |
|
scientific name |
Buthacus yotvatensis Levy et al., 1973 |
| status |
stat. nov. |
Buthacus yotvatensis Levy et al., 1973 View in CoL , stat. rev.
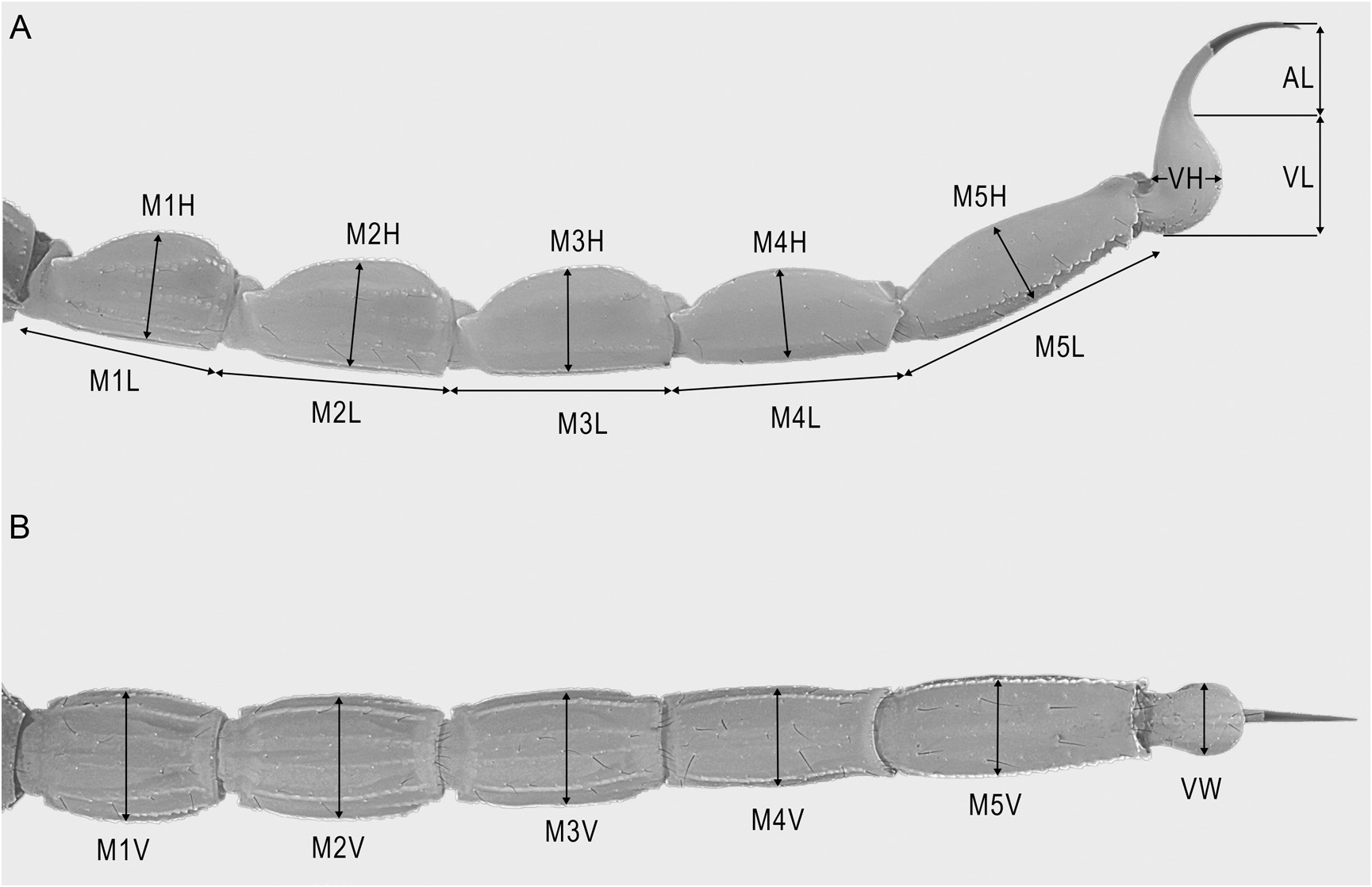
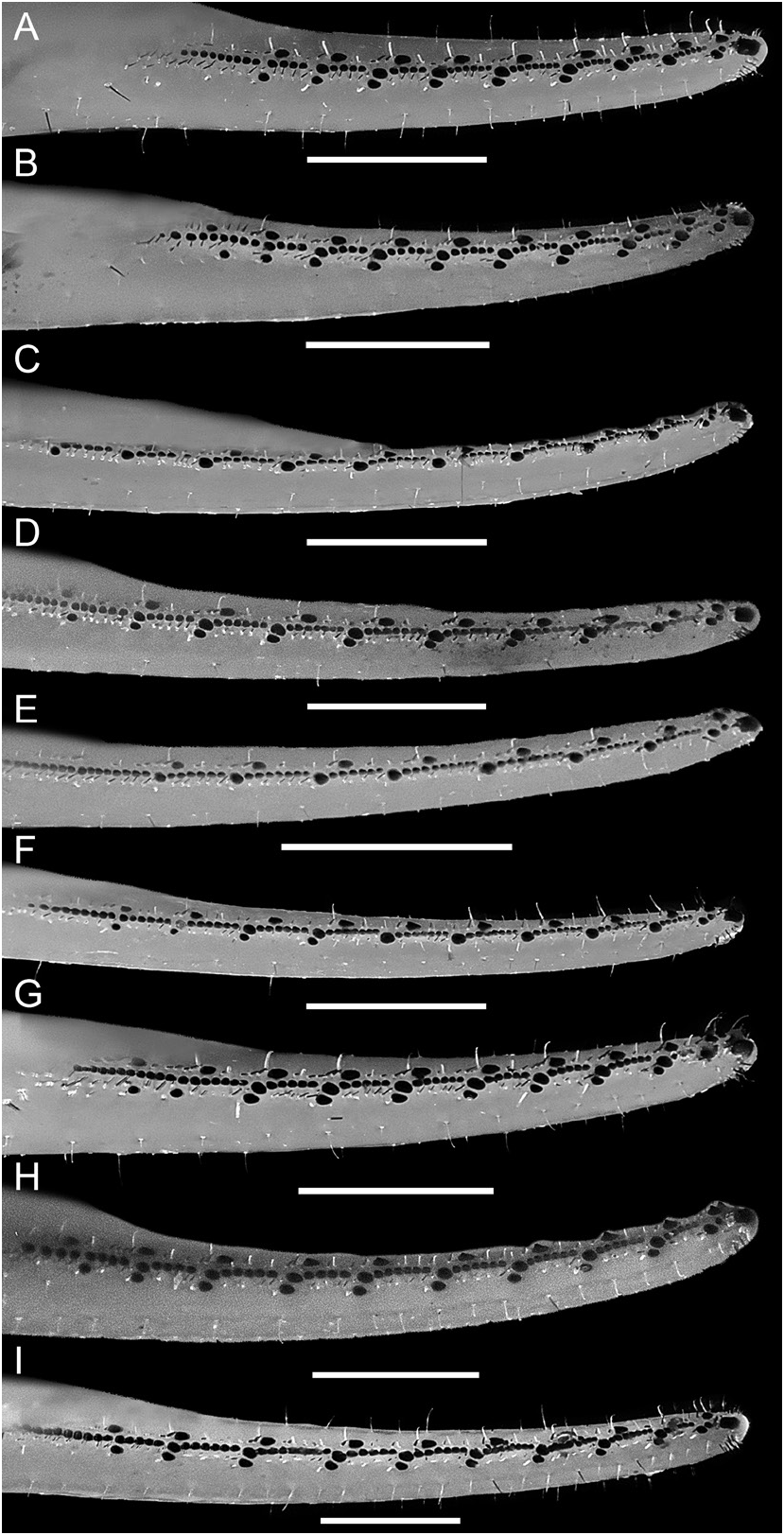
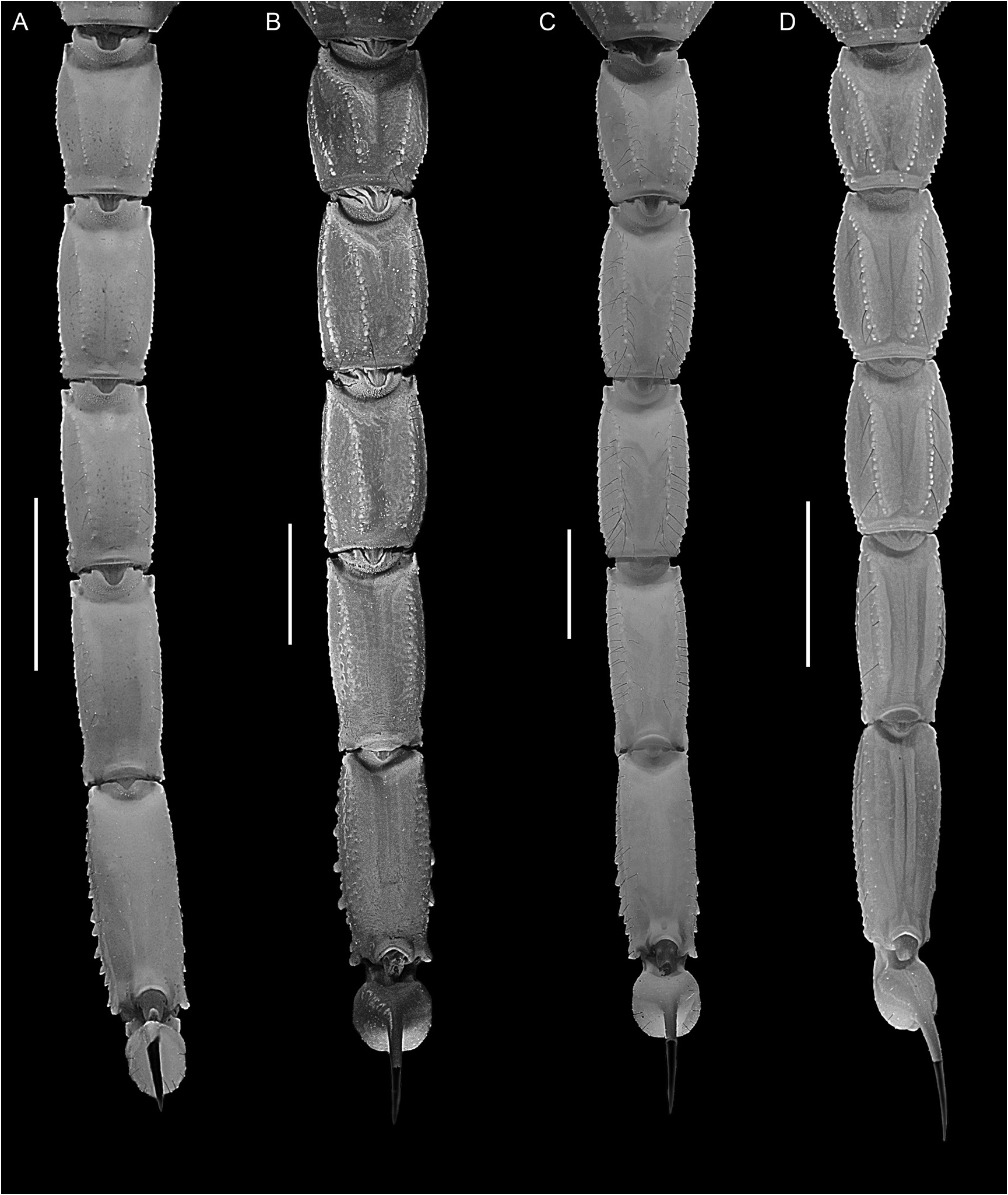
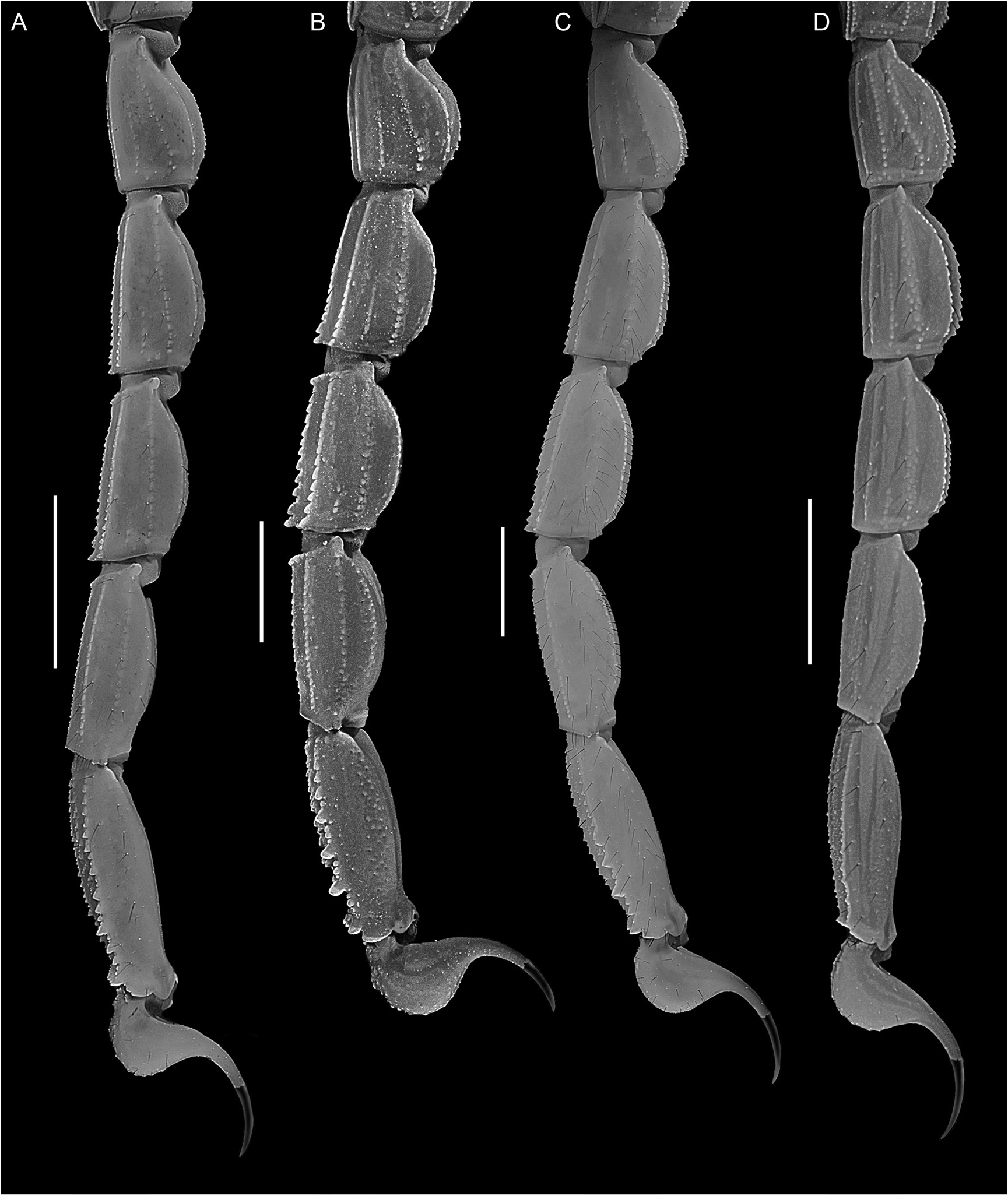
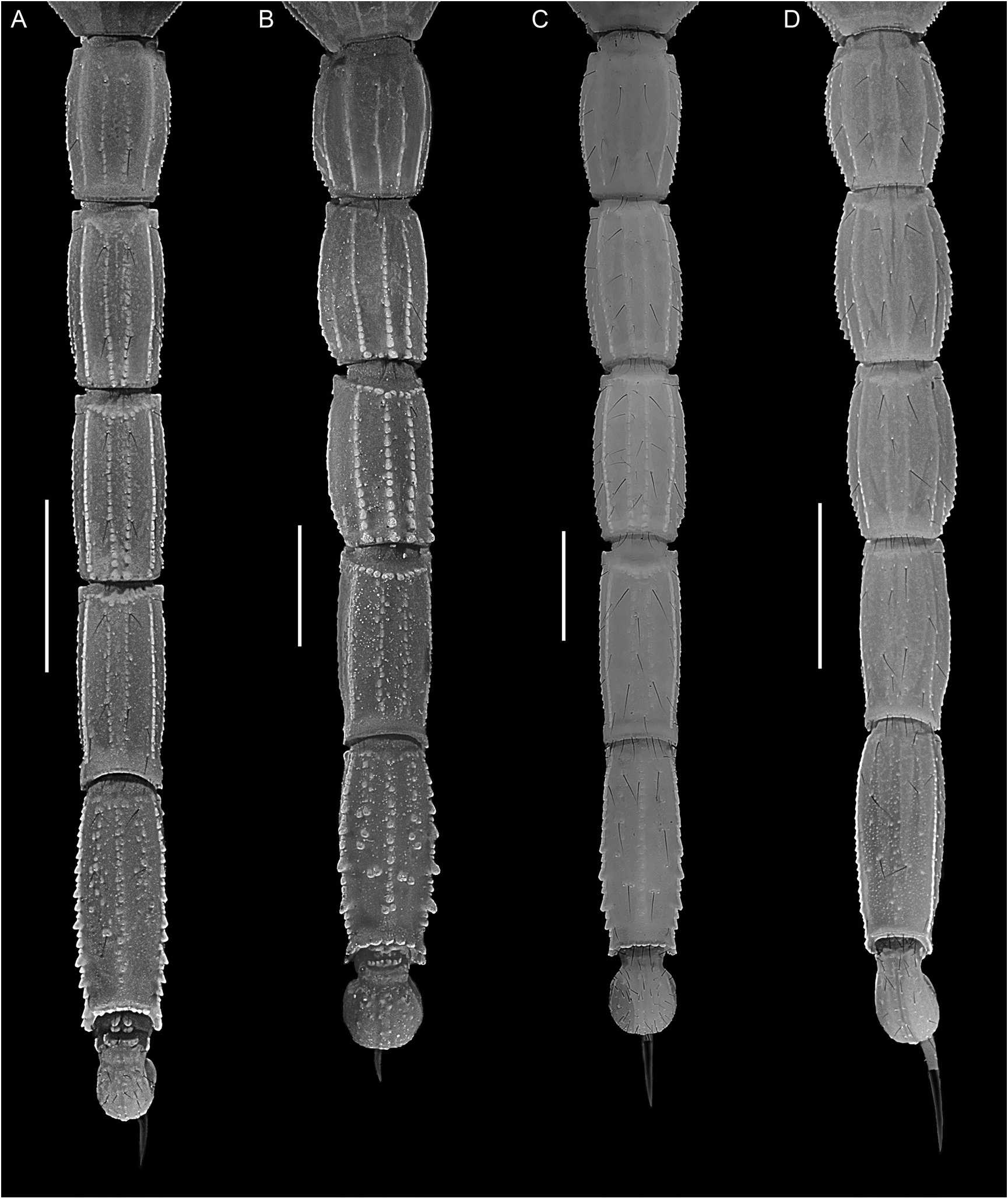
Figures 1E View FIGURE 1 , 3B, C View FIGURE 3 , 5 View FIGURE 5 , 12B View FIGURE 12 , 15 View FIGURE 15 , 16E View FIGURE 16 , F, 19E, F View FIGURE 19 , 21I View FIGURE 21 , 22G View FIGURE 22 , 23C View FIGURE 23 , 25C View FIGURE 25 , 27C View FIGURE 27 , 29G View FIGURE 29 , 51 View FIGURE 51 , 52 View FIGURE 52 , 53 View FIGURE 53 ; tables 1, 6, 7, 9
Buthacus yotvatensis Levy et al., 1973: 130–133 View in CoL , 134 (misidentification, part), figs. 32–37; Levy and Amitai, 1980: 90–93, figs. 82–85, map 6; Kinzelbach, 1984: 99, 101 (misidentification); Fet et al., 1998: 615, 616; Fet and Lowe, 2000: 85; Soleglad and Fet, 2003 a: 5 (misidentification); 2003b: 7, 149, 151, 152, 156, fig. B1-2 (misidentification); Fet et al., 2003: 3, 4, table 1 (misidentification); Hendrixson, 2006: 47, 52, 54, 56; Amr et al., 2015: 30–33, fig. 1D; Alqahtani et al., 2019: 19, 22, 24, 25, fig. 2C (misidentification); Alqahtani and Badry, 2021: 4, 10, table 1; Amr et al., 2021: 86, 88, fig. 3A, table 4; Kachel et al., 2021: 2, 3.
Buthacus yotvatensis yotvatensis: Vachon, 1979: 36 View in CoL ; Fet and Lowe, 2000: 85; Lourenço, 2004a: 206; Hendrixson, 2006: 55.
Buthacus tadmorensis yotvatensis: Kinzelbach, 1985 View in CoL : map III; Vachon and Kinzelbach, 1987: 101; Amr et al., 1988: 374; El-Hennawy, 1992: 101, 114 (part).
Buthacus tadmorensis: Vachon, 1952: 180 View in CoL (misidentification); El-Hennawy, 1992: 101, 104 (misidentification); Kovařík, 2001: 80.
Buthacus macrocentrus: Kovařík, 2005: 1 View Cited Treatment , 8; Lourenço, 2006: 64; Kaltsas et al., 2008: 214.
TYPE MATERIAL: Holotype 1 ♂ ( SMNH TAU AR 5223 View Materials old NS 5223 ), ISRAEL: Mehoz HaDarom (Southern District): Arava Valley , dunes E of Yotvata [29°53′N 35°04′E], 2.v.1965, H. Zinner [examined]. GoogleMaps
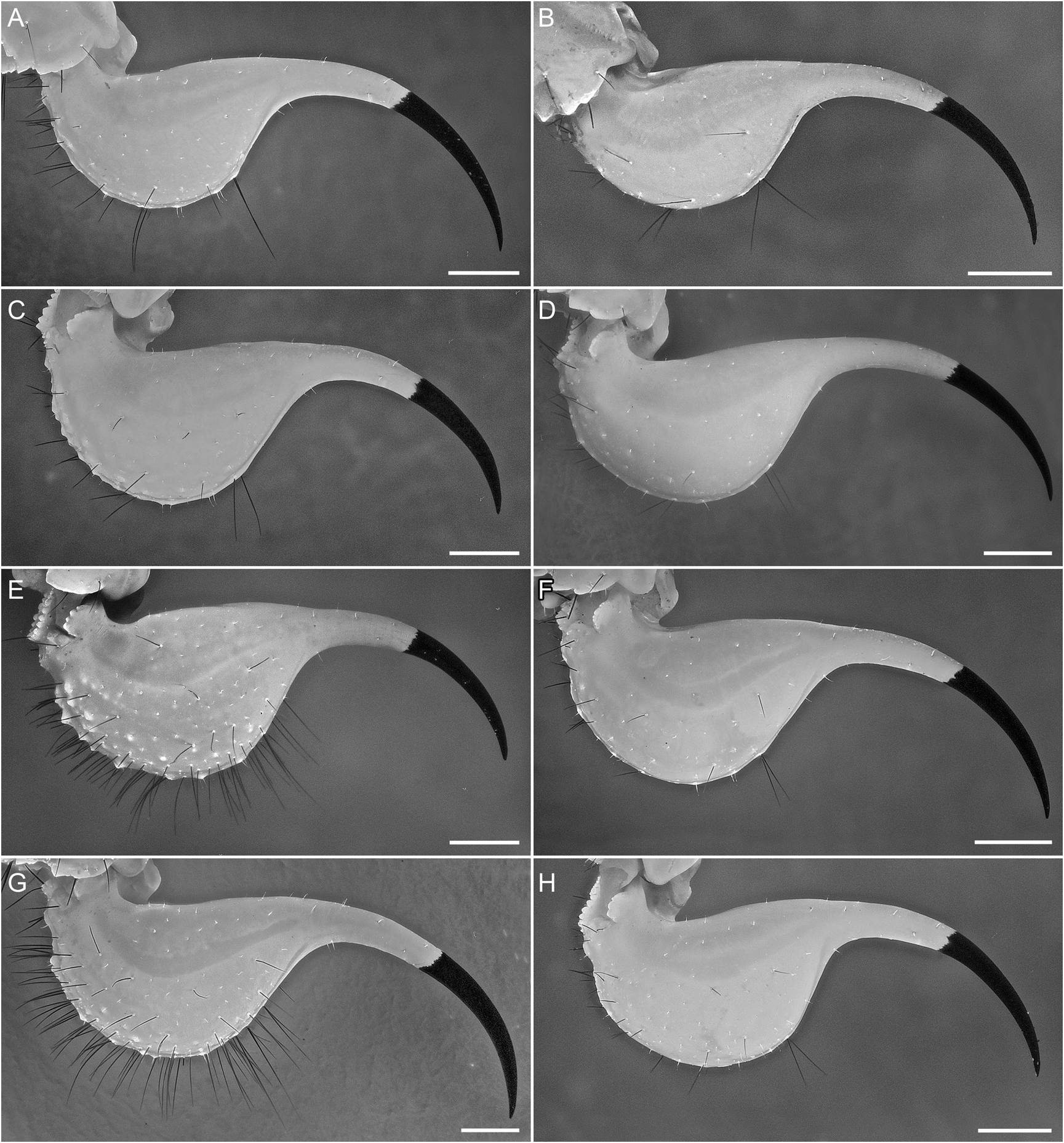
DIAGNOSIS: Buthacus yotvatensis differs from the closely related B. arava , also occurring in Israel and Jordan, as follows. The pedipalp chela of B. yotvatensis is longer and narrower, especially in the adult male (fig. 53A, B), with chela manus width:chela length, 17.4% (16.5%–18.8%, n = 8; table 6) (♂), chela manus length:movable finger length, 55.2% (51.7%–58.3%, n = 8) (♂) or 47.1% (45.5%–48%, n = 6, table 7) (♀), and chela manus width:length, 50.5% (48.1%–54.8%, n = 8) (♂) or 56.9% (51.3%–61.5%, n = 6) (♀), than that of B. arava (fig. 35A, B), with chela manus width:chela length, 29.8% (27.2%–32.8%, n = 7; table 8) (♂), chela manus length:movable finger length, 90.4% (83.5%–95%, n = 7) or 60.8% (♀), and chela manus width:length, 63.3% (57.3%– 74.5%, n = 7) or 61.6% (♀). The ungues of the leg telotarsi are shorter (approximately twothirds the length of the telotarsus) and equal on legs I and II in B. yotvatensis (fig. 22G) but longer (approximately equal to the length of the telotarsus) and unequal on legs I and II in B. arava (fig. 22C). The pectinal tooth counts are higher in B. yotvatensis , i.e., 34/35 (32–37/32–37, n = 8; table 9) (♂) and 26/26 (25–28/26–28, n = 6) (♀), than in B. arava , i.e., 20/20 (18–21/18– 22, n = 7; table 9) (♂) and 12/13 (♀). The metasomal segments and telson are densely setose in B. yotvatensis (figs. 23C, 25C, 29G), with macrosetal counts on segments I–V (sinistral/dextral), dorsosubmedian carinae, 16/15 (14–20/11–21, n = 5):25/26 (20–29/18–30):26/25 (23–31/19– 29):30/27 (24–38/23–31):12/11 (6–18/7–15), dorsolateral carinae, 9/8 (7–13/5–13):20/20 (19– 23/16–21):22/21 (16–28/18–24):15/14 (11– 20/12–17):11/13 (10–13/11–14), and telson, 102 (88–127, n = 5), but sparsely setose in B. arava (figs. 23A, 25A, 29C), with macrosetal counts on segments I–V (sinistral/dextral), dorsosubmedian carinae, 0/0 (0–1/0–1, n = 4):2/3 (2–3/2– 5):4/3 (2–5/2–6):3/3 (2–3/1–5):0/0 (0–0/0–0), dorsolateral carinae, 1/1 (0–2/0–2):2/2 (1–3/0– 3):3/2 (2–4/1–3):3/3 (2–4/2–5):5/6 (3–8/5–7), and telson, 28 (27–29, n = 4).
Buthacus yotvatensis differs from the closely related B. tadmorensis , occurring in neighboring countries, as follows. The pedipalp chela of B. yotvatensis is longer and narrower (fig. 53A, B), with chela manus width:chela length, 17.4% (16.5%–18.8%, n = 8; table 6) (♂), chela manus length:movable finger length, 55.2% (51.7%– 58.3%, n = 8) (♂) or 47.1% (45.5%–48%, n = 6; table 7) (♀), and chela manus width:length, 50.5% (48.1%–54.8%, n = 8) (♂) or 56.9% (51.3%–61.5%, n = 6) (♀), than that of B. tadmorensis (fig. 50A, B), with chela manus width:chela length, 22.3% (♂) (table 4), chela manus length:movable finger length, 64.2% (♂) or 55% (51.9%–59.8%, n = 4) (♀), and chela manus width:length, 58.7% (♂) or 59.1% (53.8%–65.1%, n = 4) (♀). The ventrosubmedian and ventrolateral carinae of metasomal segments II and III, and ventrolateral carinae of segment V are less developed in B. yotvatensis than in B. tadmorensis (fig. 25B, C), with narrow, spiniform processes posteriorly in B. yotvatensis but broad, lobate processes posteriorly in B. tadmorensis . The telson ventral surface is sparsely granular anteriorly in B. yotvatensis but densely granular anteriorly in B. tadmorensis (fig. 29E, G).
DESCRIPTION: The following description is based on specimens from across the distribution of the species in Israel (see tables 6 and 7 for measurements, and table 9 for counts).
Total length: Large scorpions, 67.4 mm (61.9– 71.9 mm, n = 8) (♂) or 76.6 mm (71.6–85 mm, n = 6) (♀).
Color: Uniformly pale yellow to yellowish olive, except as follows. Mesosomal tergites, ster- nite VII and metasomal segments I–V, each with narrow brown stripe posteriorly. Pectines pale yellow. Leg articulation points brown.
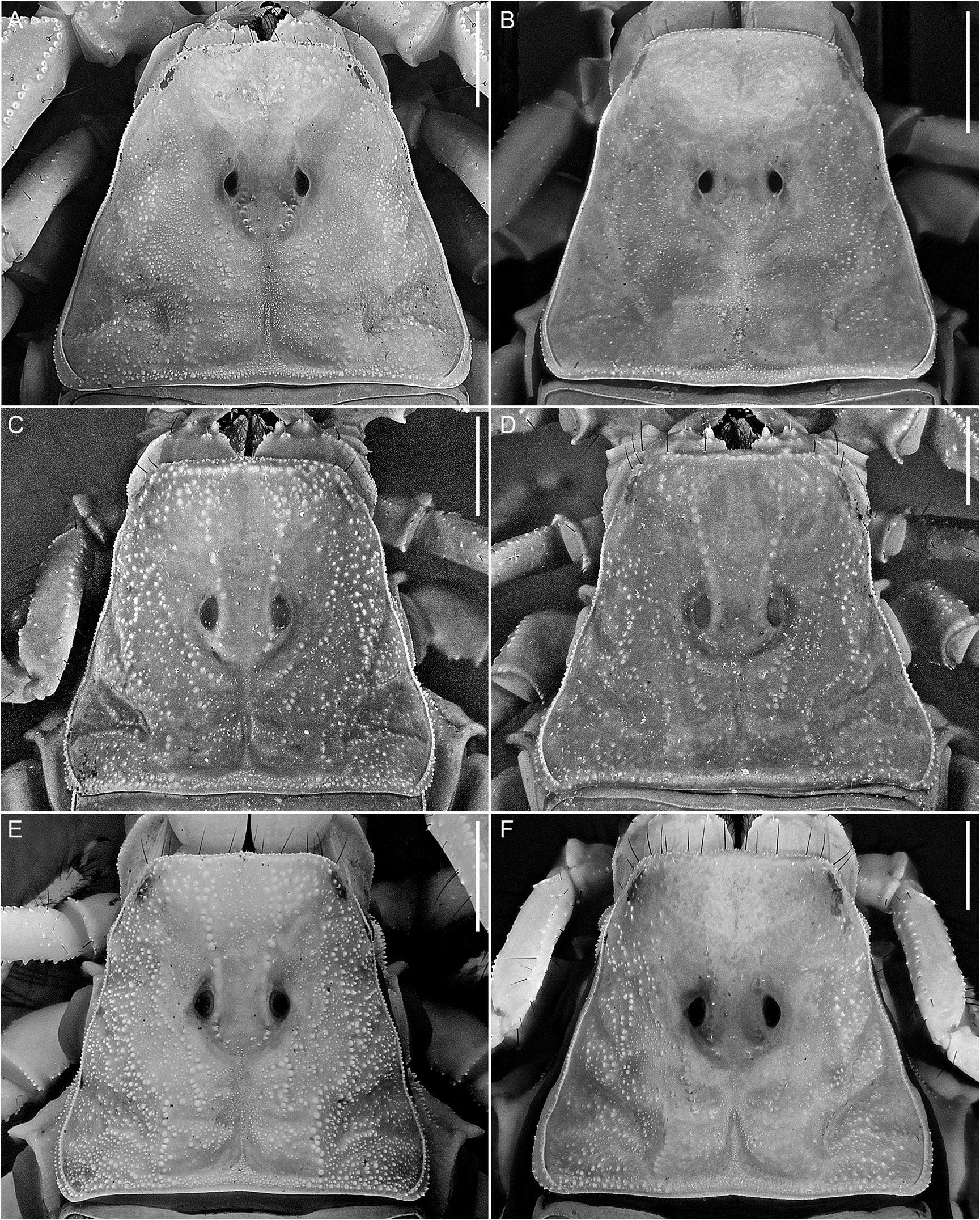
Carapace: Carapace shape trapezoidal (fig. 16C, D), anterior width:posterior width, 53.3% (46.3%–59.2%, n = 14), length:posterior width, 89.4% (81.7%–98.4%, n = 14). Five, rarely four, pairs of lateral ocelli; each lateral ocular tubercle with three major ocelli (ALMa, MLMa, PLMa), similar in size, situated anterolaterally, and one or, usually, two minor ocelli (ADMi, PDMi; PDMi may be absent) situated posterodorsal to posterior major ocellus. Median ocelli larger than lateral ocelli, distance between them more than 2× ocellus width. Median ocular tubercle situated anteromedially, distance from anterior carapace margin:carapace length, 45.4% (42.6%– 47%, n = 8) (♂) or 45.5% (43.9%–46.8%, n = 6) (♀). Superciliary and central median carinae distinct, costate-granular and weakly connected to disconnected. Anteromedian sulcus distinct; posteromedian sulcus deep, narrow anteriorly, wide posteriorly; posterolateral sulcus deep, wide, curved. Carapace intercarinal surfaces coarsely and densely granular.
Chelicerae: Cheliceral manus prodorsal margin finely granular; retrodorsal surfaces smooth or finely granular; prolateral and ventral surfaces setose. Fixed finger dorsal surface setose; dorsal margin with subdistal, medial, and proximal denticles; ventral margin with proximal and medial denticles. Movable finger smooth and glabrous; dorsal margin with retrodistal, subdistal, medial, and pair of proximal denticles; ventral margin with prodistal, medial, and proximal denticles.
Pedipalps: Femur dorsal prolateral, dorsal retrolateral and ventral prolateral carinae complete, costate-granular; prolateral ventral and prolateral ventrosubmedian carinae each comprising discontinuous row of spiniform granules; retrolateral dorsosubmedian carina obsolete, comprising discontinuous row of spiniform granules and more than 15 macrosetae; dorsal, prolateral and ventral intercarinal surfaces finely granular; retrolateral intercarinal surfaces smooth (fig. 52A, B). Patella prolateral median and ventral prolateral carinae eral (E) aspects. Scale bar = 2 mm.
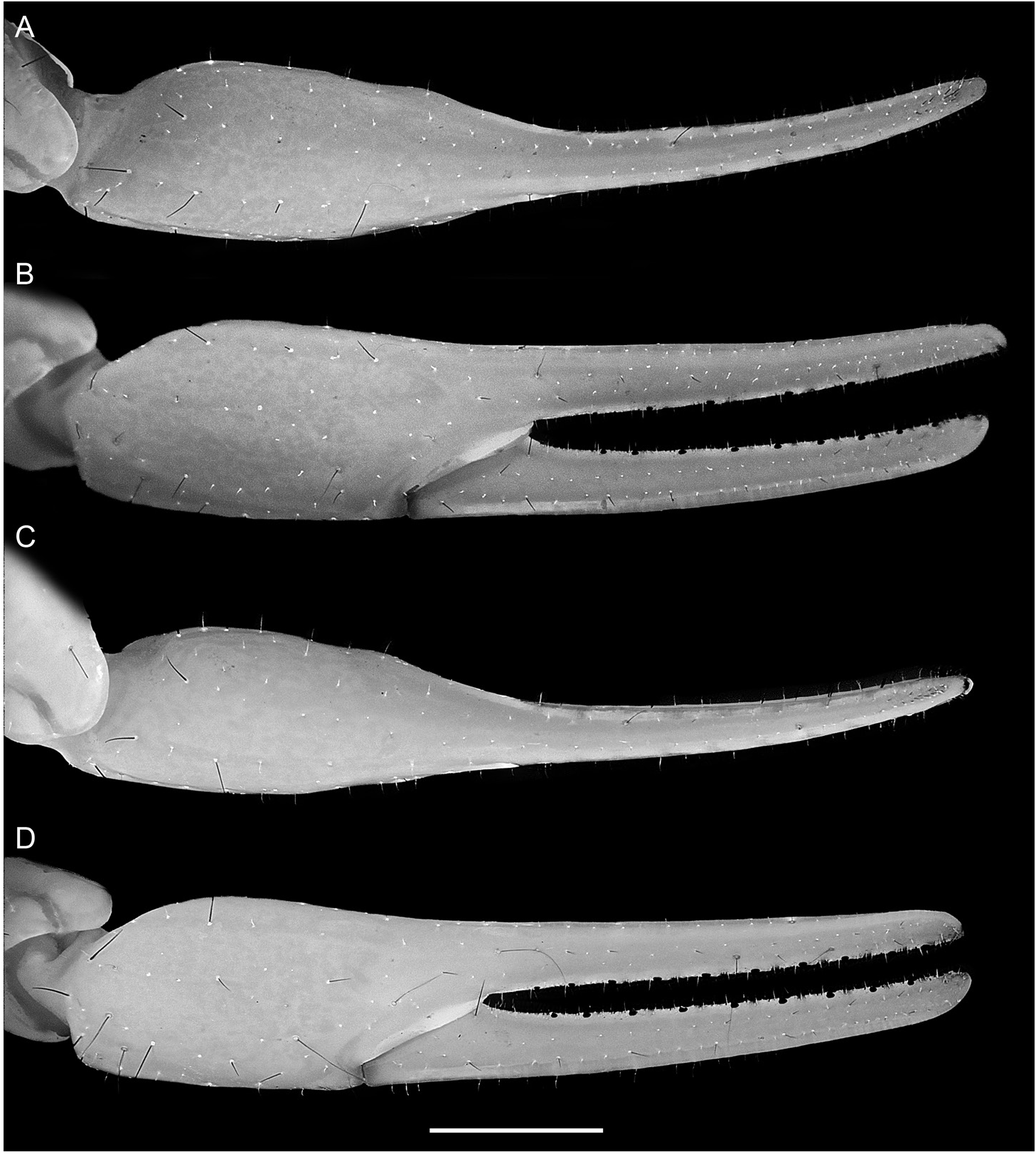
complete, costate-granular to finely serrate; dorsal prolateral carinae complete, costate-granular; other carinae obsolete; intercarinal surfaces smooth (fig. 52C–E). Chela long and slender in both sexes, manus width:length, 50.5% (48.1%– 54.8%, n = 8) (♂) or 56.9% (51.3%–61.5%, n = 6) (♀), manus height:length, 56.9% (53.3%–60.7%, n = 8) (♂) or 63.5% (60.1%–67.2%, n = 6) (♀), and manus length:movable finger length, 55.2% (51.7%–58.3%, n = 8) (♂) or 47.1% (45.5%–48%, n = 6) (♀). Chela manus acarinate; intercarinal surfaces smooth and setose (fig. 53). Fixed and movable fingers each with 9–12 (n = 12) oblique median denticle subrows; movable finger with 6–11 (n = 12) retrolateral accessory denticles (fig. 21I); proximal dentate margins of fingers straight (fig. 53B), such that no gap evident proximally when fingers closed.
Legs: Legs I–IV, femoral ventral carinae granular; patellar ventral carinae obsolete; intercarinal surfaces smooth. Legs I–IV, tibial spurs absent on I and II, present on III and IV; pro- and retroventral basitarsal (pedal) spurs present, more developed on III and IV. Legs I–IV, macrosetal counts on retrolateral margins of tibiae, 11:14:12:9; basitarsi, 13:20:23:10; telotarsi, 7:8:7:6 (n = 1). Telotarsi ungues slightly elongated, more than three-quarters of telotarsus length, equal on legs I–IV (fig. 22G).
Genital operculum: Genital opercula suboval, completely divided longitudinally, without overlapping, rounded margins (♂) or partially fused longitudinally (♀) (fig. 19E, F). Genital papillae present (♂) or absent (♀).
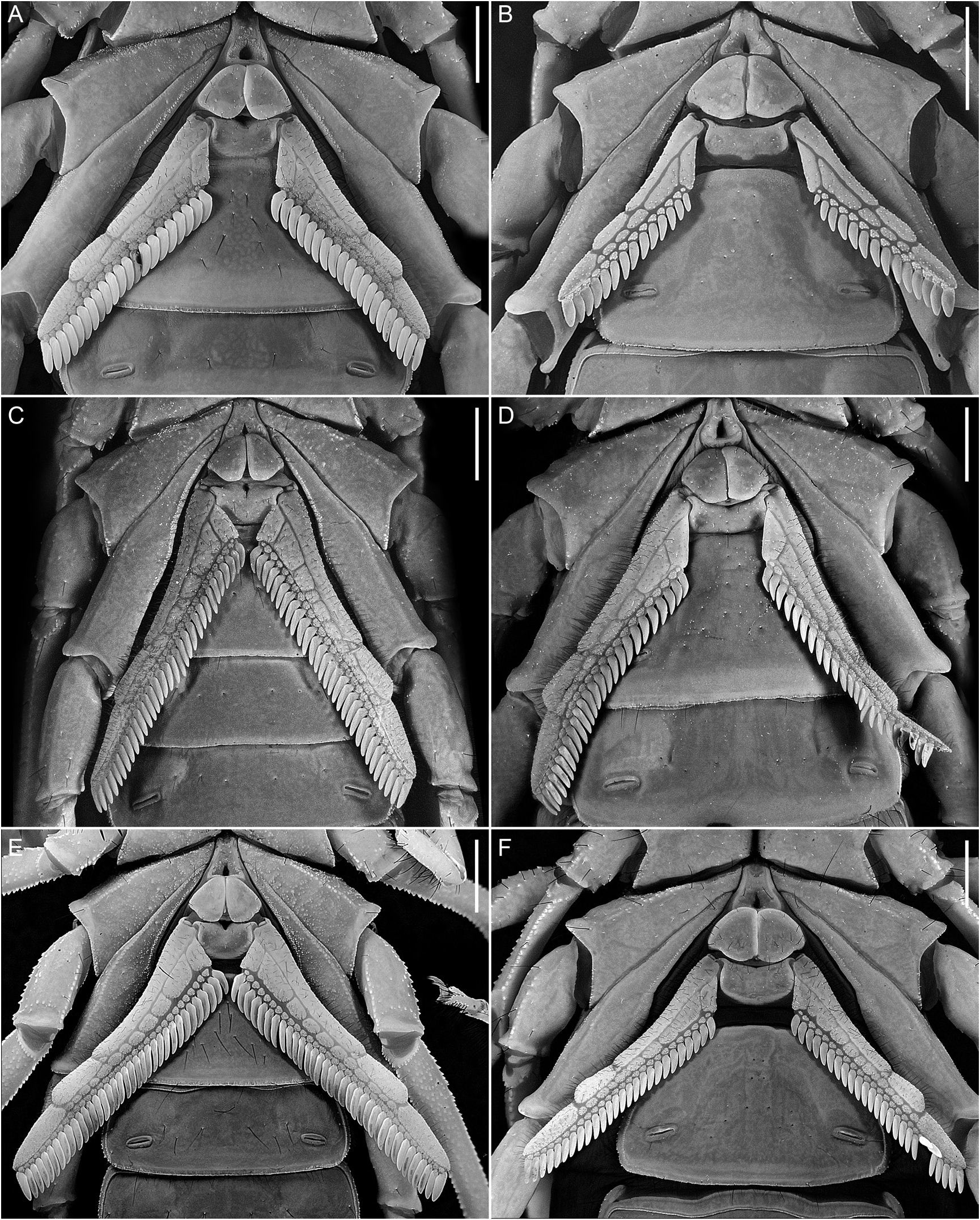
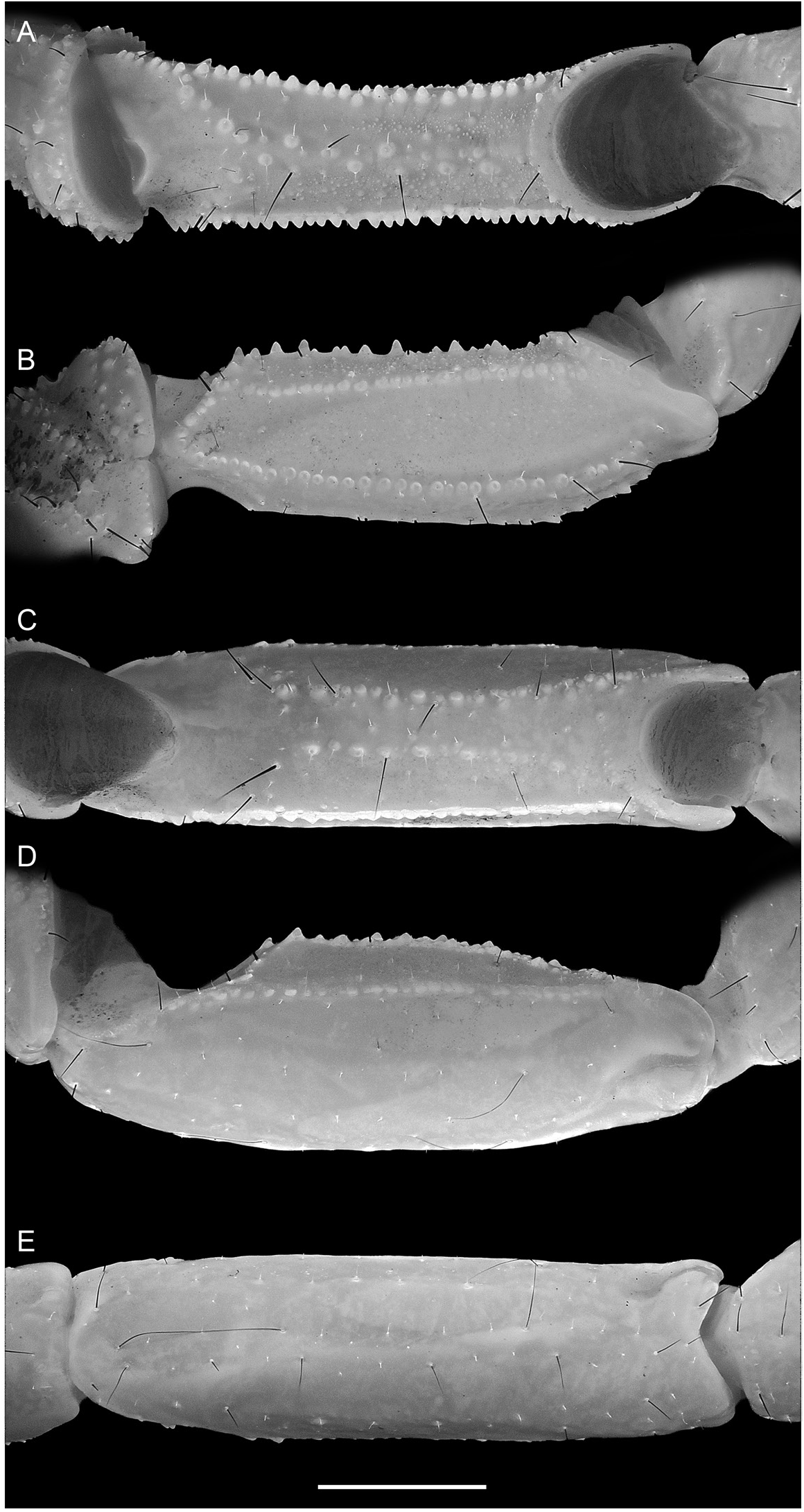
Pectines: Three marginal lamellae; 11–12 (n = 3) (♂) or 9–12 (n = 2) (♀) median lamellae (fig. 19E, F). Fulcra present. Pectinal teeth along most (♂) or part (♀) of length, dentate margin length:pecten length, 94.1% (90.4%–99.6%, n = 8) (♂) or 86.3% (81.2%–90.2%, n = 6) (♀). Pectinal teeth curved, similar in size; tooth count (sinistral/dextral), 34/35 (32–37/32–37, n = 8) (♂) or 26/27 (25–28/26–28, n = 6) (♀).
Mesosoma: Tergites I –VII progressively increasing in length posteriorly, tergite VI length:tergite VII length, 53.8% (47.7%–58.3%, n = 8) (♂) or 55.9% (51.8%–60.5%, n = 6) (♀); increasing in width posteriorly from I–IV, decreasing in width posteriorly from V–VII. Pretergites smooth; posttergites I–VI, intercarinal surfaces uniformly finely granular, becoming more coarsely and densely granular posteriorly, VII, finely to coarsely and sparsely granular. Tergites I–VI, dorsomedian carinae granular, vestigial, restricted to posterior fifth of I–III and posterior third of IV–VI; dorsosubmedian carinae granular, vestigial, restricted to posterior fifth of I–III and posterior third of IV–VI. Tergite VII, dorsomedian carina granular, vestigial, restricted to anterior half, dorsosubmedian and dorsolateral carinae distinct, granular. Sternites III–VII acarinate, smooth, and glabrous; IV–VI, respiratory spiracles (stigmata) width approximately 3× length.
Metasoma: Metasomal segments I–V becoming longer and narrower posteriorly (figs. 23C, 25C, 27C), segment I shortest, length I:II, 88% (84.3%–89.7%, n = 8) (♂) or 89.5% (87.1%–91%, n = 6) (♀); segments II–IV similar, length II:III, 96.3% (93.4%–98.1%, n = 8) (♂) or 96.7% (92.8%–100.7%, n = 6) (♀), length III:IV, 96.3% (94.1%–98.2%, n = 8) (♂) or 96.3% (93.8%–99.1%, n = 6) (♀); segment V longest, length IV:V, 86.7% (83.6%–91.8%, n = 8) (♂) or 82.3% (79.1%–85.9%, n = 6) (♀); width:length segment I, 61.1% (57.8%– 64.4%, n = 8) (♂) or 62.5% (60.7%–65.2%, n = 6) (♀), II, 51.3% (47.2%–53.7%, n = 8) (♂) or 53.5% (51.1%–56.1%, n = 6) (♀), III, 47.7% (43.2%–51%, n = 8) (♂) or 50% (48.4%–53%, n = 6) (♀), IV, 40.1% (36.3%–41.8%, n = 8) (♂) or 41.7% (39.9%–45.1%, n = 6) (♀), V, 33% (31.2%–36.2%, n = 8) (♂) or 33.8% (32.7%–35.5%, n = 6) (♀). Dorsosubmedian and dorsolateral carinae distinct, granular on segments I–III, obsolete on IV, absent on V; dorsosubmedian and dorsolateral carinae densely setose, macrosetal counts on segments I–V (sinistral/dextral), dorsosubmedian carinae, 16/15 (14/11–20/21, n = 5):25/26 (20/18– 29/30):26/25 (23/19–31/29):30/27 (24/23– 38/31):12/11 (6/7–18/15), dorsolateral carinae, 9/8 (7/5–13/13, n = 5):20/20 (19/16–23/21):22/21 (16/18–28/24):15/14 (11/12–20/17):11/13 (10/11– 13/14). Median lateral carinae distinct, granular in posterior three-quarters of segment I, posterior half of II and posterior third of III, absent on IV and V. Ventrolateral carinae distinct, granular, restricted to posterior edge of segment I; granular, with granules becoming progressively larger and subspiniform posteriorly, on II and III; costategranular on IV; serrate, comprising spiniform granules of variable size, becoming more prominent posteriorly, on V. Ventrosubmedian carinae distinct, costate on segment I; granular, with granules becoming progressively larger and subspiniform posteriorly, on II and III; costate-granular on IV; granular, restricted to anterior three-quarters of V. Ventromedian carina granular, distinct along entire length of V. Dorsal and lateral intercarinal surfaces smooth to finely and sparsely granular on segment I, smooth on II–V; ventral intercarinal surfaces smooth on I and III, finely granular across entire surface on IV and V.
Telson: Telson vesicle width:metasomal segment V width, 80.7% (74.8%–84.6%, n = 8) (♂) or 83% (77.6%–88.5%, n = 6) (♀). Vesicle globose, dorsal surface flat, ventral surface convex and rounded; height:length, 61.2% (51.6%–69.4%, n = 8) (♂) or 62.9% (59.5%–67.6%, n = 6) (♀); dorsal surface smooth; ventral surface finely granular anteriorly, smooth posteriorly; lateral and ventral surfaces densely setose, with 98 (91–102, n = 3) (♂) or 107 (88–127, n = 2) (♀) macrosetae. Aculeus long, gently curved; aculeus length:telson length, 49.6% (43.3%–55.5%, n = 8) (♂) or 51.8% (47.8%–55.5%, n = 6) (♀).
Sexual dimorphism: Adult males and females differ as follows. Males are slightly smaller, on average 67.4 mm in total length, than females, on average 76.6 mm. The carapace intercarinal surfaces are more densely granular and the carinae of the pedipalp femur and patella more coarsely granular in the male than in the female. The pedipalp chela of the male has proportionally shorter fingers than that of the female (fig. 53), as indicated by the higher chela manus length:movable finger length ratio in the male (55.2%) compared with the female (47.1%). The genital opercula are completely divided longitu- dinally, with overlapping, rounded margins in the male but partially fused longitudinally in the female (fig. 19E, F) and genital papillae are present in the male but absent in the female. The pectinal tooth count is higher in the male (32– 37) than in the female (25–28). The spiniform granules of the ventrosubmedian and ventrolateral carinae of metasomal segments II and III, and the ventrolateral carinae of segment V are less prominent in the male than in the female.
DISTRIBUTION: Buthacus yotvatensis appears to be endemic to Israel and Jordan. Most of the known records occur in the northern and southern parts of the Arava Valley, straddling the southern border between Israel and Jordan, but the species has also been recorded at one locality in the Negev (fig. 5). Whereas records from the Arava Valley range from - 294 m to 127 m in elevation, the Negev locality, Ein Zik, is situated at 345 m.
Buthacus yotvatensis is sympatric with the closely related B. arava , in the Arava Valley (fig. 4), but allopatric with B. tadmorensis , distributed from central Jordan, through Syria, southeastern Turkey and Iraq, to the northern shore of the Persian Gulf, in southern Iran (fig. 10).
ECOLOGY: Specimens of B. yotvatensis were collected at night with UV light detection on gravel flats and sparsely vegetated, stable to vegetationless, shifting inland sand dunes. Specimens were fairly common at night, mostly sitting still or walking on the ground surface, some sitting on bushes. The habitat and habitus, notably the pale coloration, smooth tegument, obsolescence of some pedipalpal and metasomal carinae, elongation of the legs, especially legs III and IV, slight dorsoventral compression of the basitarsi of legs I–III, with comblike rows of elongated macrosetae (“sand combs”) along the retrolateral margins, elongated macrosetae on the lateral and ventral surfaces of the telotarsi, and slightly elongated telotarsal ungues, are consistent with the psammophilous ecomorphotype (Prendini, 2001).
Specimens of B. yotvatensis were collected in sympatry with B. arava . Two other buthids, Androctonus crassicauda and Orthochirus scrobiculosus , also inhabit the same area, but occur on harder substrata.
REMARKS: In the original description, Levy et al. (1973: 133), stated: “ B. yotvatensis is completely different from all scorpions so far described [and] for some time it was doubted whether it should not be placed in a genus of its own.” Kinzelbach (1985) relegated B. yotvatensis to a subspecies of B. tadmorensis , a decision accepted by some authors (e.g., Vachon and Kinzelbach, 1987; Amr et al., 1988; El-Hennawy, 1992) and rejected by others (e.g., Fet and Lowe, 2000; Lourenço, 2004a). Kovařík (2005) subsequently revalidated B. macrocentrus , originally synonymized with B. leptochelys by Kraepelin (1891), and synonymized B. tadmorensis and B. yotvatensis with it. After comparing a paratype of B. yotvatensis , deposited at the MNHN, with two specimens from Palmyra, the type locality of B. tadmorensis , presumed to be part of the original type series, Lourenço (2006) suggested that B. tadmorensis is a valid species and B. yotvatensis could be a junior synonym of B. macrocentrus . Nevertheless, Kovařík et al. (2016) and others ( Kaltsas et al., 2008; Navidpour et al., 2008, 2013; Navidpour, 2015) continued to regard B. tadmorensis , B. pietschmanni , and B. yotvatensis as junior synonyms of B. macrocentrus . Lowe et al. (2019) recently rejected Lourenço’s (2006) assessment of the validity of B. tadmorensis .
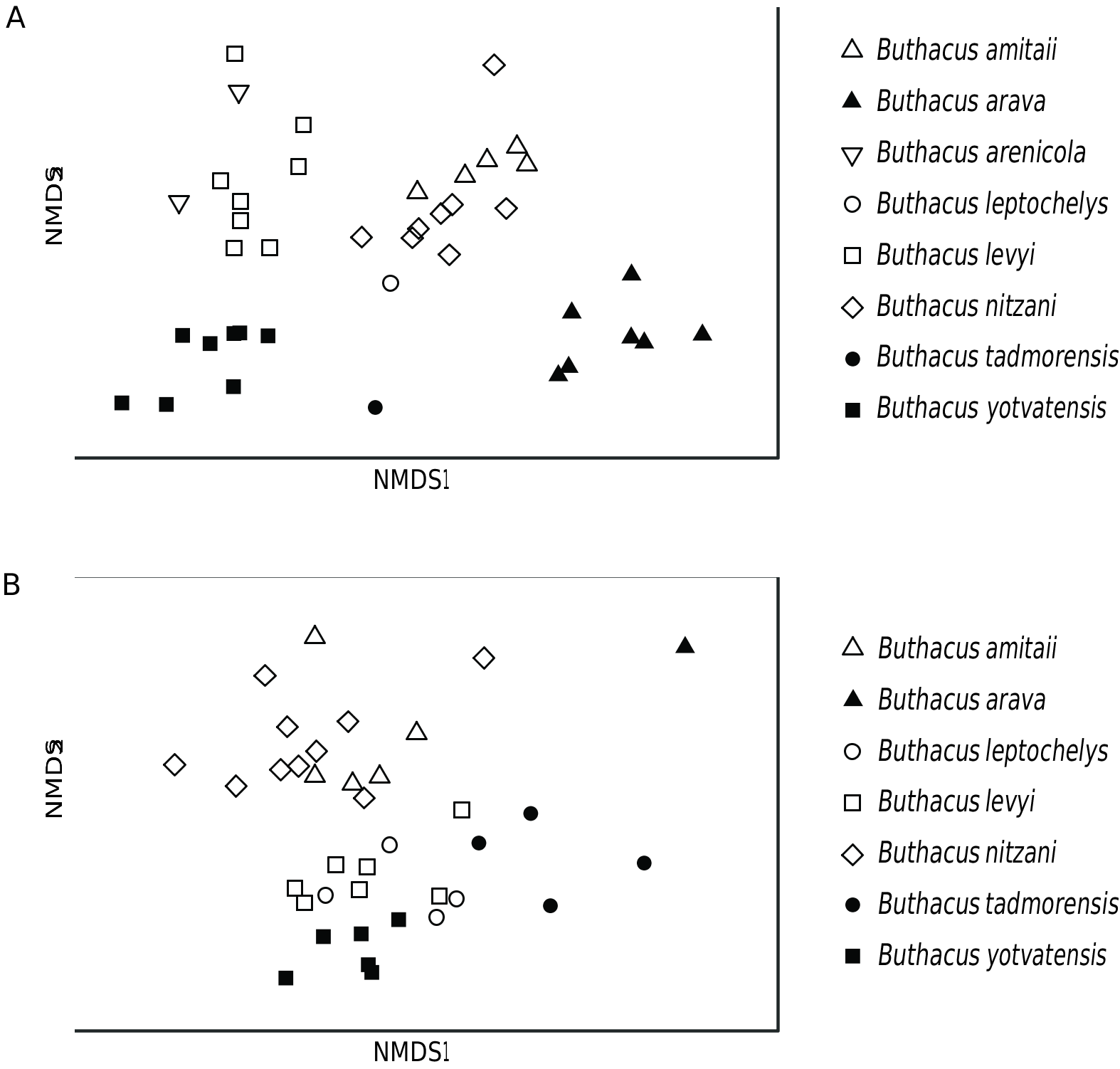
During the present investigation, examination of the types of B. leptochelys and B. macrocentrus , as well as additional material from Sinai and Israel, demonstrated that they are conspecific, as originally concluded by Kraepelin (1891), requiring B. macrocentrus to be returned to synonymy with B. leptochelys (see above). On the contrary, examination of the holotype of B. yotvatensis as well as material from across the distributions of both species, multivariate analysis of morphometrics (fig. 15), and multilocus phylogenetic analysis (fig. 14), confirmed that B. yotvatensis is a valid species, as originally indicated by Levy et al. (1973). Buthacus yotvatensis Levy et al., 1973 , stat. rev., is revalidated accordingly.
MATERIAL EXAMINED: ISRAEL: Mehoz HaDarom (Southern District): Arava Valley, Avrona Nature Reserve , 29°41′12.5″N 34°59′46.2″E, 48 m, 14–18.ix.2017, E. Gavish-Regev, N. Segev and S. Cain, 13 ♂ ( HUJ INVSC 3360 , 3362–3367 , 3370 , 3371 , 3373 , 3374 , 3376 , AMNH ex HUJ INVSC 3379 ), 1 ♀ ( HUJ INVSC 3378 ), 2 subad. ♂ ( AMNH ex HUJ INVSC 3377 , HUJ INVSC 3372 ), 1 subad. ♀ ( HUJ INVSC 3368 ), 3 juv. ♂ ( HUJ INVSC 3369 , 3375 , 3380 ), 1 juv. ♀ ( HUJ INVSC 3361 ) GoogleMaps ; Ein-Zik campsite, 30°48′22.4″N 34°51′03.1″E, 345 m, 11.ix.2018, Y. Zvik, 2 ♂ ( HUJ INVSC 3483 , 3485 ), ♀ ( HUJ INVSC 3484 ), juv. ♀ ( HUJ INVSC 3481 , 3482 ) GoogleMaps ; Hai-Bar Yotvata Nature Reserve , 29°50′51.0″N 35°01′42.5″E, 68 m, 26.viii.2011, L. Prendini and T. L. Bird, 32 ♂, 2 ♀, 1 subad. ♀ ( AMNH), 1 juv. ♂, 3 juv. ♀ ( AMCC [ LP 11172 ]), 4.viii.2016, E. Gefen and S. Cain, 3 ♂ ( AMCC [ LP 15075 ] ex HUJ INVSC 3331 , HUJ INVSC 3333 , 3335 ), 1 ♀ ( AMCC [ LP 15076 ] ex HUJ INVSC 3332 ), 2 subad. ♀ ( HUJ INVSC 3329 , 3334 ), 1 juv. ♂ ( HUJ INVSC 3330 ) GoogleMaps ; Hazeva [30°42′N 35°15′E], - 129 m, 7.vii.1983, A. Bouskila, 1 ♀ ( HUJ INVSC 2532 ) GoogleMaps ; Hazeva, Nahal Gidron , 30°46′52.2″N 35°14′33.5″E, - 136 m, 16. ix.2017, Y. Olek and S. Cain, 1 ♂ ( HUJ INVSC 3352 ) GoogleMaps ; Hazeva, Nahal Mashaq , 30°47′26.1″N 35°15′26.5″E, - 155 m, 16.ix.2017, Y. Olek and S. Cain, 2 ♂ ( AMNH ex HUJ INVSC 3354 , HUJ INVSC 3355 ), 3 ♀ ( AMNH ex HUJ INVSC 3353 , 3357 , HUJ INVSC 3356 ) GoogleMaps ; Hazeva, Nahal Shezaf , 30°44′36.0″N 35°16′03.2″E, - 134 m, 16.ix.2017, Y. Olek and S. Cain, 1 ♂ ( AMCC [ LP 15080 ] ex HUJ INVSC 3359 ), 1 subad. ♀ ( HUJ INVSC 3358 ) GoogleMaps ; Iddan, N of ( Hamada ), 30°50′N 35°16′E, - 174 m, 15.ix.1988, B. Shalmon, 1 subad. ♂ ( HUJ INVSC 2611 ) GoogleMaps ; Lotan , SE of Lotan Plantations, 30°00′10.4″N 35°05′44.1″E, 127 m, 15.ix.2017, Y. Olek and S. Cain, 5 ♂ ( AMCC [ LP 15079 ] ex HUJ INVSC 3350 , HUJ INVSC 3346 , 3347 , 3349 , 3351 ), 1 ♀ ( AMCC [ LP 15078 ] ex HUJ INVSC 3348 ) GoogleMaps ; Nahal Sha’alav , 4 km S of Kibbutz Yahel, 30°02′40.6″N 35°06′33.7″E, 153 m, 15.ix.2017, Y. Olek and S. Cain, 3 ♂ ( AMCC [ LP 15077 ] ex HUJ INVSC 3342 , AMNH ex HUJ INVSC 3343 , HUJ INVSC 3344 ), 1 ♀ ( HUJ INVSC 3345 ) GoogleMaps ; Qetura [29°58′N 35°03′E], 100 m, M. Israel, 8.x.1974, 1 ♂ ( HUJ INVSC 2330 ), 20.x.1974,1 ♀ ( HUJ INVSC 2332 ), 17.iv.1975, 2 ♀ ( HUJ INVSC 2336 , 2337 ) GoogleMaps ; Samar [29°50′N 35°01′E], 103 m, 25.ii.1985, U. Mendel, 1 subad. ♀ ( HUJ INVSC 2612 ) GoogleMaps ; Samar sand dunes, E of Eliphaz date plantations, 29°48′06.6″N 35°02′19.2″E, 64 m, 4. viii.2016, E. Gefen and S. Cain, 4 ♂ ( AMCC [ LP 15073 , 15074 View Materials ] ex HUJ INVSC 3311 , 3312 , HUJ INVSC 3313 , 3314 ), 30.vii.2017, S. Cain, 4 ♂ ( AMNH ex HUJ INVSC 3319 , 3320 , HUJ INVSC 3316 , 3317 ), 1 ♀ ( HUJ INVSC 3318 ), 24.v.2017, S. Cain, 1 subad. ♀ ( HUJ INVSC 3315 ) GoogleMaps ; Samar sand dunes, old quarry, 29°48′25.9″N 35°02′06.6″E, 70 m, 13.ix.2017, Y. Olek and S. Cain, 3 ♂ ( AMNH ex HUJ INVSC 3308 , HUJ INVSC 3305 , 3307 ), 1 subad. ♀ ( HUJ INVSC 3306 ) GoogleMaps ; Samar sand dunes, E side of Samar date plantations, 29°48′44.2″N 35°02′27.9″E, 67 m, 4.viii.2016, E. Gefen and S. Cain, 1 ♂ ( HUJ INVSC 3321 ), 24.v.2017, S. Cain, 4 ♂ ( AMNH ex HUJ INVSC 3324 , 3327 , HUJ INVSC 3322 , 3326 ), 1 ♀ ( AMNH ex HUJ INVSC 3323 ), 1 subad. ♂ ( AMNH ex HUJ INVSC 3328 ), 1 juv. ♂ ( HUJ INVSC 3325 ) GoogleMaps ; Samar sand dunes, N part of Samar Nature Reserve , 29°49′36.8″N 35°02′35.8″E, 65 m, 13.ix.2017, Y. Olek and S. Cain, 5 ♂ ( AMNH ex HUJ INVSC 3303 , 3304 , HUJ INVSC 3300–3302 ) GoogleMaps ; Samar sand dunes, Samar Nature Reserve , 29°49′06.9″N 35°02′40.2″E, 64 m, 13.ix.2017, Y. Olek and S. Cain, 2 ♂ ( AMNH ex HUJ INVSC 3309 , HUJ INVSC 3310 ) GoogleMaps ; Timna , sand dunes E of, [29°45′N 35°00′E], 109 m, 24.iii.1972, Y. Ayal, 1 ♂ ( HUJ INVSC 2303 ), 1 juv. ♀ ( HUJ INVSC 2304 ), 22.ii.1972, M. Broza, 1 juv. ♀ ( HUJ INVSC 2305 ) GoogleMaps ; Wadi Fukra [Zin Tichon, 30°50′N 34°49′E], 384 m, 20.iii.1944, 1 ♀ ( HUJ INVSC 2492 ) GoogleMaps ; Yotvata , [29°53′N 35°03′E], 64 m, 20.x.1974, M. Israel, 1 ♀ ( HUJ INVSC 2333 ) GoogleMaps ; Yotvata , E side of circle field, 29°53′28.7″N 35°04′40.5″E, 72 m, 14. ix.2017, Y. Olek and S. Cain, 3 ♂ ( AMNH ex HUJ INVSC 3340 , HUJ INVSC 3339 , 3341 ) GoogleMaps ; Yotvata , NE side of circle field, 29°54′02.6″N 35°04′42.1″E, 77 m, 30.vii.2017, S. Cain, 2 ♂ ( AMNH ex HUJ INVSC 3337 , HUJ INVSC 3338 ) GoogleMaps ; Yotvata, SE side of circle field, 29°53′21.8″N 35°04′40.9″E, 71 m, 30.vii.2017, S. Cain, 1 ♂ ( AMNH ex HUJ INVSC 3336 ). JORDAN: Karak Governorate: Wadi Khanzeerah (Khanzairh), 30°53′39.9″N 35°25′38.3″E, - 294 m, 9.ix.2013, L. Prendini, Z. Amr and L. Al Azam, 1 ♀ ( AMNH). Tafilah Governorate: Wadi Al Ghwaibeh (Ghwaqibeh), 30°47′35.6″N 35°23′39.7″E, - 119 m, 9. ix.2013, L. Prendini, Z. Amr and L. Al Azam, 2 ♂, 1 ♀ ( AMNH), [leg] ( AMCC [ LP 13561 ]) GoogleMaps ; Wadi Al Ghwaibeh ( Ghwaqibeh ), 30°48′13.3″N 35°24′21.6″E, - 106 m, 9.ix.2013, L. Prendini, Z. Amr, and L. Al Azam, 2 ♂, 1 ♀ ( AMNH) GoogleMaps .





No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Buthacus yotvatensis Levy et al., 1973
| Cain, Shlomo, Gefen, Eran & Prendini, Lorenzo 2021 |
Buthacus macrocentrus: Kovařík, 2005: 1
| Kaltsas, D. & I. Stathi & V. Fet 2008: 214 |
| Kovarik, F. 2005: 1 |
Buthacus yotvatensis yotvatensis:
| Hendrixson & B. E 2006: 55 |
| Lourenco, W. R. 2004: 206 |
| Fet, V. & G. Lowe 2000: 85 |
Buthacus tadmorensis yotvatensis:
| El-Hennawy, H. K. 1992: 101 |
Buthacus tadmorensis:
| Kovarik, F. 2001: 80 |
| El-Hennawy, H. K. 1992: 101 |
Buthacus yotvatensis
| Amr, Z. S. & M. A. A. Baker & M. Al-Saraireh & D. A. Warrell 2021: 86 |
| Kachel, H. S. & Al-Khazali, A. M. & Hussen, F. S. & Yagmur, E. A. 2021: 2 |
| Amr, Z. & O. A. Abed & T. Al-Share & N. Hamidan & L. Prendini 2015: 30 |
| Hendrixson & B. E 2006: 47 |
| Fet, V., B. & Gantenbein, A. & Gromov, G. & W. R. Lourenco 2003: 3 |
| Fet, V. & G. Lowe 2000: 85 |
| Fet, V., G. A. & W. D. Sissom 1998: 615 |
| Kinzelbach 1984: 99 |
| Levy, G. & P. Amitai 1980: 90 |
| Levy, G. & P. Amitai & A. Shulov 1973: 133 |