
Archaeochionelasmus nekvasilovae Kočí, Newman & Buckeridge, 2017
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4294.2.3 |
|
publication LSID |
lsid:zoobank.org:pub:2D289A54-7600-4677-B487-84F91A4719CA |
|
DOI |
https://doi.org/10.5281/zenodo.6023946 |
|
persistent identifier |
https://treatment.plazi.org/id/2F651BDE-7839-4995-A1F1-2A81D2B45D5C |
|
taxon LSID |
lsid:zoobank.org:act:2F651BDE-7839-4995-A1F1-2A81D2B45D5C |
|
treatment provided by |
Plazi |
|
scientific name |
Archaeochionelasmus nekvasilovae Kočí, Newman & Buckeridge |
| status |
sp. nov. |
† Archaeochionelasmus nekvasilovae Kočí, Newman & Buckeridge , sp. nov.
Text-fig. 6; Pls. 1, 2
urn:lsid:zoobank.org:pub: 177CC951-CEC1-425C-B693-46ADB83BD5D9
Diagnosis. As for the genus.
Etymology. Named in honour of Dr Olga Nekvasilová (*1928), a prominent palaeontologist who contributed greatly to the knowledge of Mesozoic brachiopods as well as the Late Cretaceous hard-substrate communities. Dr Nekvasilová discovered this specimen in 1963.
Holotype. NM-O7615, collections of the National Museum , Prague.
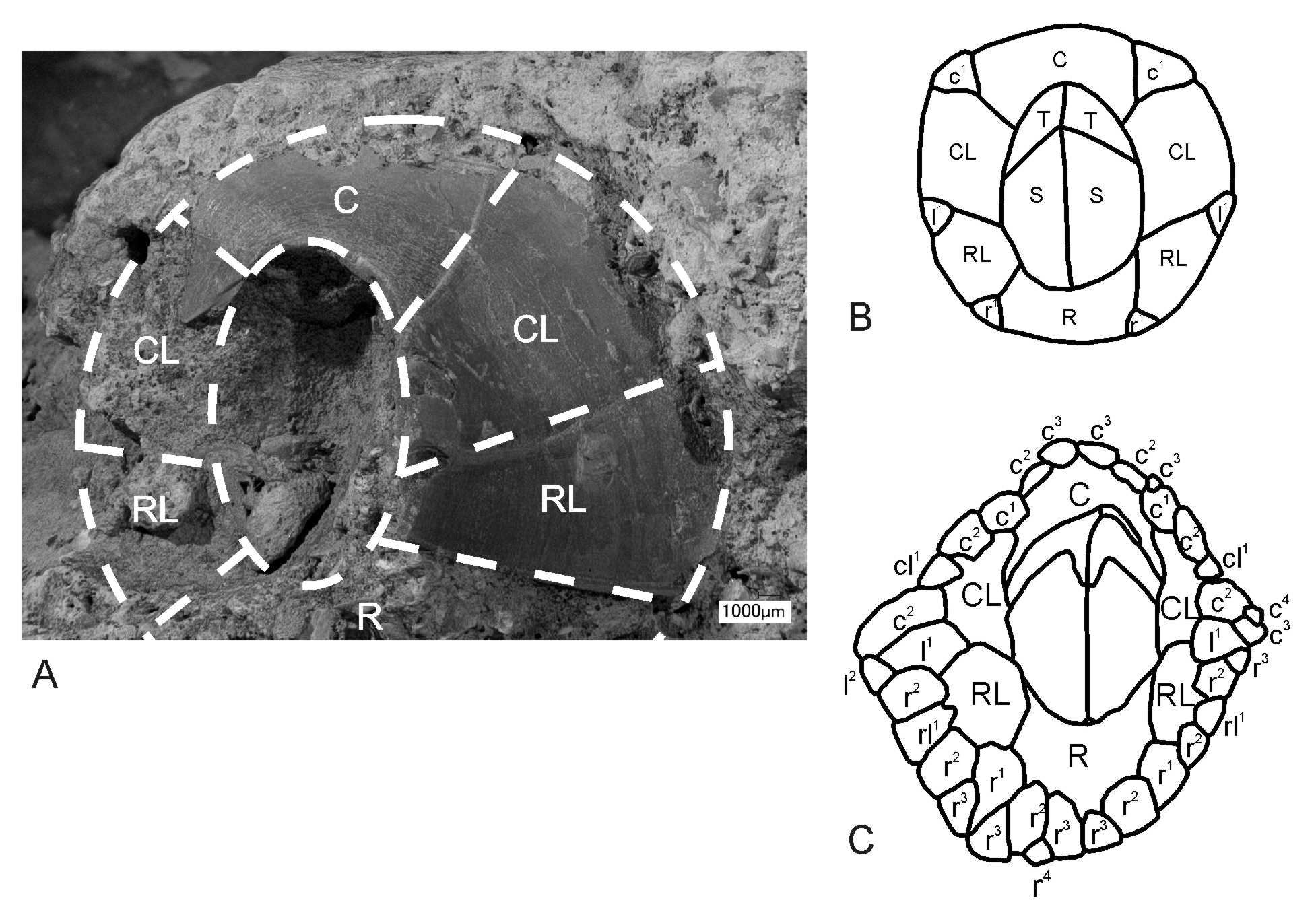
Material. Three articulated parietal plates—carina, left carinal latus and left rostral latus. The form of the rostrum is readily reconstructed, but the nature of the operculum and IPs is unknown.
PLATE 1. A–C General views of the specimen. Rostrolatera (RL) is oriented down and to the left in A and B; rightmost in C is carina. D Detail of base of carinolatera (CL) with possible traces of original imbricating plates. Note the rough surface observable at CL base to left (see this feature also in Pl. 2G). E Detailed view of junction of CL and C plates at their basalmost portion. F Detailed view of ala of C. A–C are identical in scale, A–C digital macrophotography, D–E optical microphotography, F 3D microscope image.
PLATE 2. A–E detailed views of rostrolatera (left)/carinolatera (right) junction. A–B Top-lateral views, C Top view. D–E Details in similar view using refracted light. Although these plates show partial recrystallization, some internal features are still observable in refracted light. F Detail of RL/CL junction showing overlapping margin of RL. G Detail of the RL/CL junction at their basal portions with rough surface. H Detail showing left radius of the RL plate. Compare delicate ornamentation of the plate surface with F. D–E are identical in scale. A is a 3D microscope image, B–H optical microphotography.
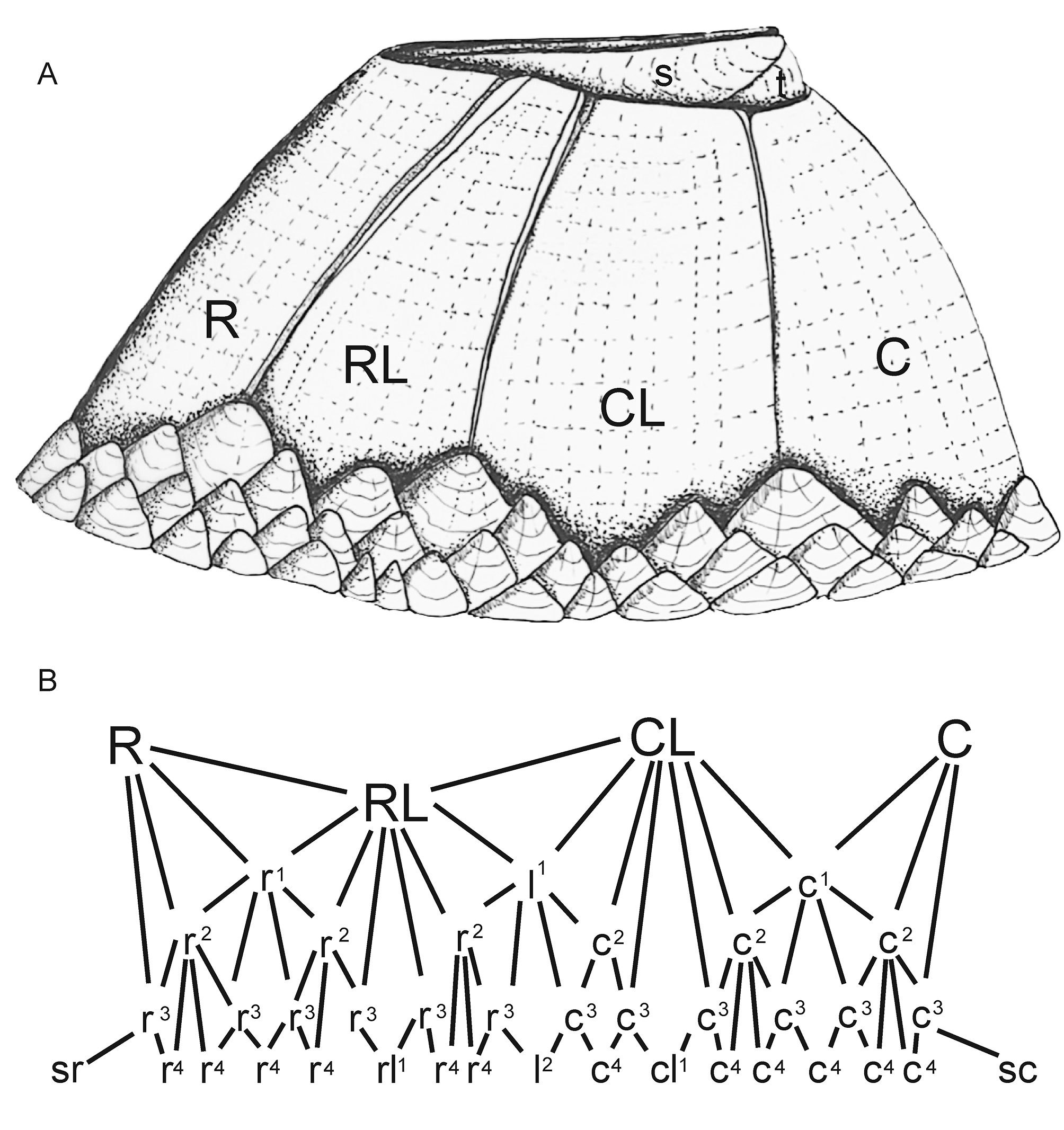
Description. Shell smooth, medium-low conical, comprising six compartmental plates—C, CL1, RL, R (rostrum reconstructed). Pseudo-radii narrow, running sub-parallel to the articular margins, slightly overlapping the edges of the parietes along both margins of the RL and the carinal margin of the CL1 ( Fig. 3 View FIGURE 3 , 6 View FIGURE 6 , 7), separated from them by a well-formed longitudinal groove; wider nearer summit, with width increasing by accretion of the fine transverse and longitudinal ridges; the transverse growth lines are not upturned (as in true radii that develop later in balanomorph evolution). Note that these pseudo-radii are not precursors of the true radii, which developed later in the Palaeogene. The longitudinal ridges represent successive growth increments, with new ridges and grooves gradually added at the base; RL and CL1 rhomboid, with summits parallel to the base; C semi-conic. RL—narrow: 16 mm basal length: 9.9 mm, length in upper portion 2.4 mm; CL1—high 9– 5 mm, length in basis 13 mm, length in upper portion 7.7 mm.
Although no complete whorls of IPs are retained, the basal margin of the parietal plates (where not eroded or damaged) and the exposed ala (Pl.1, Fig. F) have variously arranged indentations indicating where tissue within extended out underneath the basal IPs and the wall. The delicate, sinuous, lateral calcareous ridges occurring near the base of the compartments (Pl. 1, Fig. D) are interpreted as sites that entrapped fine calcareous sediment along the zone where the imbrications abutted the compartments. There are also some fragments of shell near the base that could be the remains of IPs that survived the preparation of the specimen (Pl. 1, Fig. E).
Mode of life and palaeoecology. A reconstruction of Archaeochionelasmus nov. gen. ( Fig. 6 View FIGURE 6 ) shows a moderately low profile similar to that of the “turtle barnacle” Emersonius Ross & Newman, 1967 recorded in the Eocene of Florida (USA), Protochelonibia from the Lower Miocene of the Central Paratethys ( Austria) ( Harzhauser et al. 2011), extant Chelonibia (Zardus et al. 2013) , Platylepas and Stomatolepas ( Southward 2008) . The form of Archaeochionelasmus nov. gen. is consistent with that required for attachment to living substrates such as an ammonite or turtle, both of which are recorded in the BCB (Ekrt et al., 2001; Kear et al., 2014). Although direct evidence is lacking, we suggest that Archaeochionelasmus nov. gen. may have lived as an obligate commensal with ammonites and/or marine reptiles. Notwithstanding this, it is quite possible that it lived on hard, inorganic substrates in the littoral zone. However, further indirect support for commensal life is the absence of any other neobalanomorph plates, conspecific or otherwise, at this locality. Although negative evidence, it suggests there were no other barnacles present, compared to, for example, the variety of forms known to have lived on hard substrates along the rocky coast at Ivö Klack (southern Sweden) during the Campanian ( Withers 1935; Gale & Sørensen 2015).
| BCB |
Universitat Aut�noma de Barcelona |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |