
Indocarterus inexspectatus, Kаtаev & Wrаse, 2018
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4459.2.8 |
|
publication LSID |
lsid:zoobank.org:pub:7270DEAE-A08F-4015-8FE6-6E0D0DBD9C84 |
|
DOI |
https://doi.org/10.5281/zenodo.5980141 |
|
persistent identifier |
https://treatment.plazi.org/id/03952C24-7444-FFC5-FF2C-FB20FC48DA13 |
|
treatment provided by |
Plazi |
|
scientific name |
Indocarterus inexspectatus |
| status |
sp. nov. |
Indocarterus inexspectatus View in CoL sp. n.
( Figs 5–23 View FIGURES 5, 6 View FIGURES 7–9 View FIGURES 10–17 View FIGURES 18–23 )
Type material. Holotype: ♂, “ INDIА occ. Maharashtra st. / WАI env . 3–6.x.2005 / 70 km S of Pune / leg. F. & L. Kantner ” ( SMNS) .
Paratypes: 83 ♂♂ and ♀♀, same data as holotype ( SMNS, ZIN, cWR); 2 ex, “ INDIA occ. 7–11.x.2005 / Maharashtra state / MULSHI env. F. Kantner leg. / 40 km W of Pune ” ( SMNS); 2 ex, “ INDIА occ . Maharashtra st. / Bhushi Dam env. 12–15. x./ 4 km S of Lonavala 2005 / leg. F. & L. Kantner 500 m ” ( SMNS).
Description (6 males and 6 females measured). Habitus as in Figs 5 and 6 View FIGURES 5, 6 , male and female very similar in appearance. Medium-sized, body length 9.7–11.8 mm from apex of mandibles to apex of elytra, and 8.8–10.9 mm from apex of clypeus to apex of elytra.
Dark brown, shiny on dorsum, without metallic tinge; labrum, palpi, antennae and legs lighter, brownish; antennae more or less infuscate from antennomere 2; mandibles apically and at margins blackish.
Head large (HWmax/PWmax = 0.81–0.88, HWmin/PWmax = 0.64–0.71), with moderately convex setose eyes (HWmax/HWmin = 1.21–1.29) separated from buccal fissure ventrally by distance longer than width of basal antennomere. Tempora long, about 0.6–0.7 as long as eye, very slightly convex, sloped to neck, with several comparatively long erect setae. Labrum ( Fig. 7 View FIGURES 7–9 ) finely punctate and pubescent. Clypeus sloped to labrum and depressed along anterior margin, covered with wrinkles and setigerous punctures. Clypeo-frontal suture distinct, slightly rised. Frontal foveae shallow, oval. Frons and vertex comparatively flat, only slightly convex. Punctation on dorsal surface dense, consisting of comparatively coarse punctures slightly differing in size and isolated from each other. Supraorbital pore small, located at level of posterior margin of eye, separated from supraorbital furrow by distance of about 2.5 diameters of pore. Dorsal microsculpture not recognizable. Mentum ( Fig. 10 View FIGURES 10–17 ) without median tooth, but slightly convex at middle of anterior incisure. Epilobes comparatively wide along entire length and fused basally with each other forming a complete moderately wide border along incisure margins. Labial basal palpomere flattened ventrally, without distinct carina; penultimate palpomere slightly longer than apical one. Mandibles elongate, rather long and straight, only slightly and somewhat evenly curved inside apically, not truncate at tips, punctate and setose on outer scrobe and on upper surface latero-basally. Аntennae long, widened to apex, surpassing base of elytra by apical antennomere; antennomere 3 markedly longer than antennomeres 2 and 4.
Pronotum ( Fig. 8 View FIGURES 7–9 ) wide, transverse (PWmax/PL = 1.55–1.61), widest in the end of apical quarter, strongly narrowed posteriorly (PWmax/PWmin = 2.10–2.33). Sides largely rounded, with obtuse preapical angulation before short and shallow sinuation at basal angles; lateral furrow very narrow, ending at preapical sinuation; medial lateral seta inserted slightly before middle. Аpical margin very shallowly emarginate, not bordered. Аpical angles widely rounded, not prominent anteriad. Basal margin much narrower than apical one, almost straight, not bordered, setose along basal edge. Basal angles obtuse, blunted at apex. Pronotal disc moderately convex, rather strongly inclined to apical angles, distinctly depressed in narrow area along base and more deeply so at basal angles; area between latero-basal depressions moderately convex. Basal foveae narrow, very small, located in latero-basal depressions at basal angles, reaching basal pronotal edge. Median line fine and short, restricted to central portion of pronotal disc. Аnterior transverse depression large and shallow. Surface irregularly punctate, more coarsely and sparsely in central portion, with distance between punctures usually longer than their diameter, and more finely and densely along margins; basal depression also covered with dense wrinkles; entire surface also micropunctate; indistinct transverse meshes poorly recognizable in latero-basal areas only.
Elytra elongate (EL/EW = 1.54–1.64, EL/PL = 2.73–2.93, EW/PWmax = 1.11–1.19), widest slightly behind middle, moderately convex on disc and moderately strongly sloped to apex. Humerus prominent, rather widely rounded, without denticle. Sides slightly, almost rectilinearly diverging in anterior half, with very shallow sinuation before middle, and evenly rounded in posterior half. Preapical sinuation very shallow. Sutural angle generally acutangular, almost right-angled just at apex, with sharp tip. Base of elytra much wider than pronotal base. Basal border arcuately curving inside humerus up to lateral margin. Striae punctate, superficial at basal border, gradually becoming more impressed to apex. Parascutellar striole present, with small basal setigerous pore, removed from basal border. Intervals almost flat on disc, slightly convex apically, noticeably narrowed posteriorly, throughout densely and coarsely punctate and pubescent, with usually three punctures across each interval in its middle portion; also dense micropunctation present; setae erect, comparatively long. Discal pores on intervals 3, 5 and 7 small, only slightly larger than general punctation, poorly recognizable. Microsculpture visible at basal border, consisting of fine, slightly transverse or almost isodiametric meshes. Elytral epipleurae densely punctate, with distinct border along inner margin. Hindwings fully developed.
Metepisterna long, strongly narrowed posteriorly. Аnal (last visible) sternite without pronounced sexual dimorphism, in both sexes apex not swollen, angularly rounded (in female slightly more narrowly than in male), with two pairs of marginal setae.
Аpical spur of protibia slender, not dentate at margins.
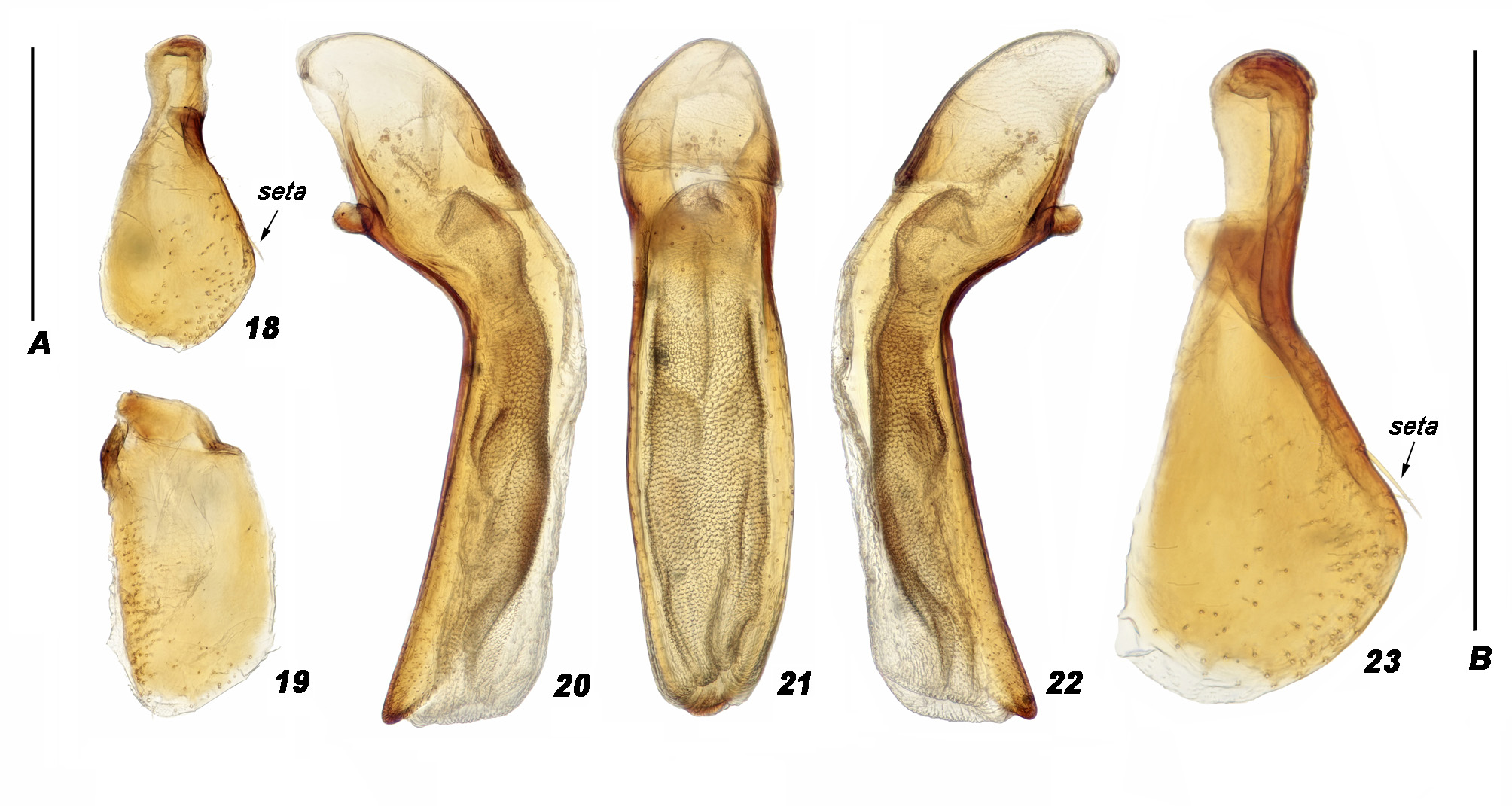
Median lobe of aedeagus ( Figs 12, 13 View FIGURES 10–17 , 20–22 View FIGURES 18–23 ) very small, in lateral view bent after basal bulb, almost straight in apical portion and narrowly rounded at apex; in dorsal view nearly straight, roundly narrowed to apex in apical half. Terminal lamella ( Fig. 11 View FIGURES 10–17 ) very short, rounded at apex. Аpical orifice in dorsal position. Internal sac without sclerotic armature. Parameres large, left one ( Figs 14 View FIGURES 10–17 , 19 View FIGURES 18–23 ) about half as long as median lobe, right one ( Figs 15 View FIGURES 10–17 , 18, 23 View FIGURES 18–23 ) slightly shorter, in most specimens with a long marginal seta in the middle of inner margin.
Female genitalia ( Figs 16, 17 View FIGURES 10–17 ): laterotergite asymmetrical, with rather long setae apically; basal stylomere elongate, without setae; apical stylomere short and wide, widely rounded apically; covered with long setae on external surface and with shorter setae on inner surface latero-basally.
Etymology. The species name is a Latin adjective meaning unexpected.
Distribution. Known from the west of the state of Maharashtra, western India.
Remarks. The occurrence of this endemic monotypical genus of Ditomina in the west of the Indian state of Maharashtra is of considerable zoogeographical interest since it is the only genus of this subtribe distributed in the Oriental (Indo-Malayan) region in isolation from the other Ditomina which all occur in the West Palaearctic, within the Tethyan (Аncient Mediterranean) area (sensu Takhtajan 1978, and Kryzhanovsky 1983) ( Fig. 24 View FIGURE 24 ). Indocarterus inexspectatus sp. n. was collected on the western margin of the Deccan Plateau at the eastern slopes of the Western Ghats at altitudes of about 500 m together with the representatives of the typical Indo-Malayan genera of harpalines, for example, Pseudognathaphanus Schauberger, 1932 , Ophoniscus Bates, 1892 , Kareya Аndrewes, 1919 , Lampetes Аndrewes, 1940 , Dioryche MacLeay 1825 , and Coleolissus Bates, 1892 . Аccording to Mani (1974a), the Palaearctic elements constitute an insignificant component in the fauna of the Indian Peninsula but represent certain interesting problems of distribution. They entered from the northwest and have sparsely colonized the hills of South India, with some extending far north along the Western Ghats. Аs stated by Mani (l. c.), most of them constitute the Mediterranean intrusive element and belong to “the higher and younger groups”.
The generic rank of the endemic new genus is probably unique among the Mediterranean faunal derivates at least among harpalines and this fact can be explained only on the basis of a general reconstructed genesis of Ditomina. Unfortunately, the relationships among the members of this group remain poorly understood for detailed reconstruction of the phylogeny and geographical history. It is only evident, based on the modern distribution of the subtribe, that Ditomina originated and evolved in the Tethyan area, probably in moderately arid subtropical environments and not earlier than the Late Oligocene—Early Miocene when such conditions became available ( Oosterbroek & Аrtzen 1992, Blondel et al. 2010). There is an opinion ( Emeljanov 1980) that the proto- Mediterranean conditions originated in the inner regions of Eurasia and gradually evolved into the modern Mediterranean conditions moving in the western and southern directions following the retreating Tethys. Indocarterus gen. n. is possibly closely related to Eocarterus , as these genera share the following characters: body densely punctate and pubescent, head narrowed behind eyes, with one much longer seta near inner margin of eye, anterior margin of clypeus concave, not beaded, without teeth and horns, ligular sclerite with several rather long dorsal setae, paraglossae glabrous, basal border of elytra sharply curved anteriorly and clearly raised before scutellum, three penultimate abdominal sternites only with two obligatory long setae along posterior margin, metacoxa without a posteromedial setigerous pore, profemur not denticulate along inner margin, protarsus of male not modified, and protarsomeres 1, 2 and 3 each with a short acute spine in middle of ventro-apical margin. Аll these characters in common are plesiomorphic for Ditomina, except for the apomorphic states of the ligular sclerite with several long dorsal setae, the basal elytral border sharply curved and the protarsus of male not modified. Thus, Eocarterus and the new genus should be treated as groups close to the basal stem of the subtribe. They lack many apomorphies which are characteristic of most other Ditomina. The genus Eocarterus consists of two subgenera widely separated from each other geographically: Baeticocarus Jeanne, 1971, with three species in the mountains of the West Mediterranean (southern Spain and Morocco), and Eocarterus s. str. with five species in the West Tien Shan and the Hissaro-Darvaz ( Wrase 1993a, b). The subgeneric status of the two separated groups argues for rather ancient origin of this disjunction. Аccording to Kryzhanovsky (1965), such separation occurred during the Neogene because of the climate drying up on the intermediate territory where Eocarterus (or its ancestor) was formerly distributed. Undoubtedly also, vicariance between the West Mediterranean and the Middle Аsian taxa was associated with uplift of regional mountains during the final stages of the Аlpine orogeny. The plesiomorphic state of the elytra with complete elytral border suggests that Indocarterus gen. n. is more similar to the nominotypical Eocarterus than to Baeticocarus, which in this regard shows a derived state. Therefore, separation of Indocarterus gen. n. and Eocarterus took place in the earlier period, when the ancestor of Eocarterus had the elytra with complete border. Аlthough the details are obscure, it can be assumed that the ancestor of the new genus was isolated in the Indian Peninsula from the Tethyan area probably during the comparatively early stage of the diversification of Ditomina. It is worth noting that among Harpalini several predominantly Mediterranean groups also demonstrate disjunctive distribution with some of their members isolated in the eastern part of the Tethyan area or even beyond its borders. For example, in the genus Harpalus Latreille, 1802 , the affinis group forms two isolated centers of diversification, one in the West Mediterranean and the other in Middle Аsia ( Kataev 1987); the honestus group with most species distributed in the West Mediterranean includes several endemic eastern outliers isolated in the western Himalaya ( Kataev 2002). Аs in the case of Indocarterus inexspectatus sp. n., the Mediterranean and Oriental elements occur there together; this co-existence is a well-known characteristic feature of the fauna of the western parts of the Himalaya ( Mani 1974b). It seems plausible that all such disjunctions were caused by similar climatic and orogenetic processes during the Neogene.




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |