
Caddo dentipalpus ( Koch & Berendt, 1854 )
|
publication ID |
https://doi.org/10.3897/zookeys.16.224 |
|
publication LSID |
lsid:zoobank.org:pub:DB5973A9-8CF6-400B-87C4-7A4521BD3117 |
|
DOI |
https://doi.org/10.5281/zenodo.3791612 |
|
persistent identifier |
https://treatment.plazi.org/id/0392774B-785C-106E-B6DD-53E0FB82FAE6 |
|
treatment provided by |
Plazi |
|
scientific name |
Caddo dentipalpus ( Koch & Berendt, 1854 ) |
| status |
|
Caddo dentipalpus ( Koch & Berendt, 1854) View in CoL
Figs 10-14 View Figures 10-11 View Figures 12-14
Synonymy. See Dunlop (2006, p. 169).
Holotype. MfN, Berendt collection nr. 7340, redescribed by Dunlop (2006), from Baltic amber (Palaeogene, Eocene); precise locality unclear.
Additional material. MfN, MB.A. 1655 (also bears the label “Ser. 12/13”) and MB.A. 1656 (also bears the label “Ser. 8”), from Bitterfeld amber, probably from the site of the Goitsche open-cast Mine near Bitterfeld, Sachsen-Anhalt, Germany; Palaeogene (Oligocene: Chattian).
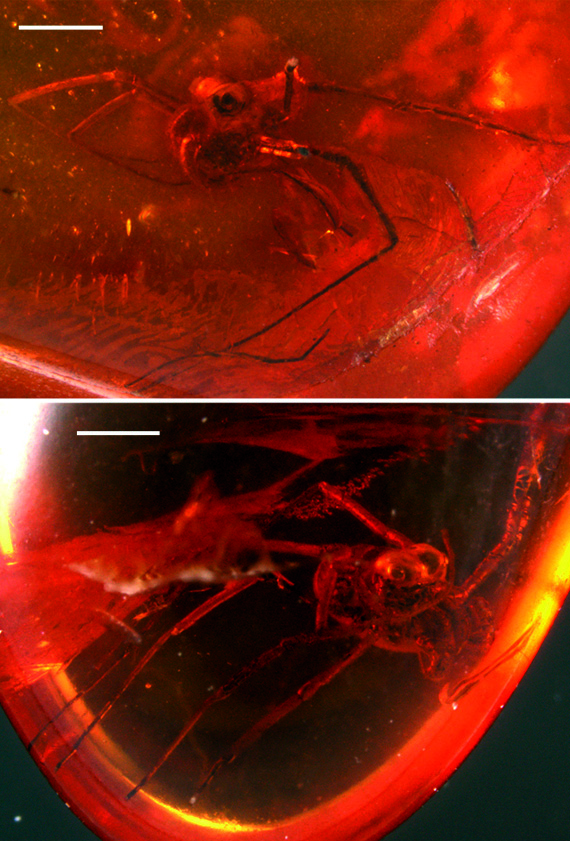
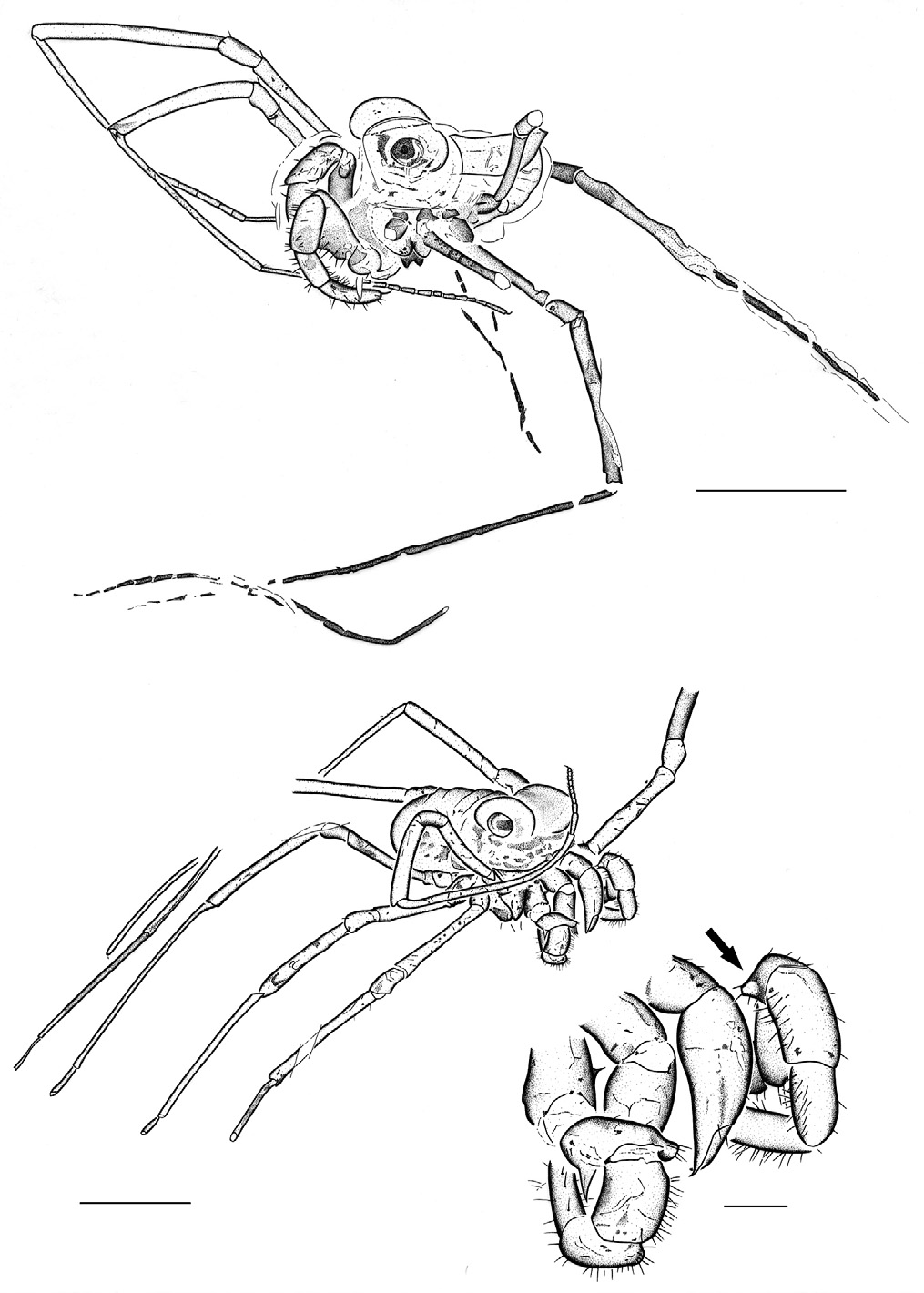
Description. MB.A. 1655 ( Figs 10 View Figures 10-11 , 12 View Figures 12-14 ) is a relatively complete specimen in lateral view. Body compact, length ca. 1.5 mm. Prosoma dominated dorsally by massive, oval eyes, maximum width 0.58 mm, in the form of a bilobed ocular tubercle. Further details of carapace morphology concealed beneath eyes. Chelicerae and sternal region cannot be seen. Pedipalps stout, femur particularly robust with at least eleven stout setae on the inferior lateral surface opposing the more distal articles. Femoral dentition equivocal in this specimen. Palpal article lengths (in mm): femur, 0.46; patella, 0.40; tibia, 0.23; tarsus 0.42. Some setal positions on femur revealed as sockets only. More distal palpal articles also setose; tarsus ends in a single, gently curving claw. All legs elongate and slender, femora and patellae sometimes with a short, stout seta near the distal end; patella noticeably shorter than adjacent articles. Right legs 1, 2 and probably 4 relatively complete. Articles of leg 2 noticeably longer than corresponding articles of leg 1, e.g. patellae 0.26. and 0.29 mm, tibiae 1.02 and 1.29 mm and metatarsi 1.36 and 1.91 mm respectively. Right leg 1 ends in a distally annulate tarsus with at least 11 annulations; distal annulus bears a single, hook-shaped claw. Left leg 3 relatively complete, other legs on left side truncated. A disarticulated leg fragment, probably a distal region bearing an annulate tarsus, crosses the distal part of left leg 3. Opisthosoma covered dorsally with a shield-like sclerite, length 0.67 mm, lacking obvious segmental divisions. Ventral region equivocal, but some hints of up to four sternites towards the posterior end of opisthosoma preserved.
MB.A. 1656 ( Figs 13-14 View Figures 12-14 ) is another excellent specimen, also in lateral view, and preserving the mouthparts in some detail. Body compact, length c. 1.5 mm. Prosoma with large eyes, maximum diameter 0.56 mm, remainder of carapace obscured. Chelicerae robust, composed of three articles. First cheliceral article tubular, total length not preserved, ornamented with sparse setae. Second cheliceral article somewhat swollen and globose proximally, tapering distally and becoming recurved at the very end to form the fixed finger of the claw (= chela). Total length 0.57 mm. Movable finger short, length 0.16, forming the free finger of the claw. Pedipalps robust. Right trochanter bears one thorn medially. One femur preserves a prominent, dentate thorn on the inferior surface (hence the name dentipalpus ) and both femora exhibit a blunt, mesal protuberance at the distal end of the article, bearing setae. Patella, tibia and tarsus of pedipalp strongly setose. Leg 1 complete, folded across the body, with podomere lengths (in mm) of femur, 0.60; patella, 0.32; tibia, 0.79; metatarsus, 1.79; and tarsus, 0.59. Total length 4.09. Tarsus multi-articulate, composed of ten elements with the longest located proximally, and ending in a single tarsal claw. Remaining legs incomplete, but longer than leg 1. Dorsal opisthosoma with evidence for at least five tergites. Ventral surface equivocal.
Remarks. Caddo dentipalpus ( Koch & Berendt, 1854) is present in both Baltic and Bitterfeld amber. It is of particular interest given that there are no Recent records of caddids in Europe, or much of Asia for that matter. According to Shear (1975), Caddidae is currently restricted to North America, Mexico, Chile, Japan, Australia, New Zealand and South Africa. Like these amber inclusions, the genus Caddo is restricted today to the northern hemisphere. The amber shows that it used to occur more widely, being present in north-central Europe during the Palaeogene, but subsequently became extinct in this region. The Bitterfeld specimens described here are probably juveniles. Their body lengths of 1.5 mm are shorter than the 2.3 mm of the holotype in Baltic amber and are slightly outside the published minimum ranges for adults of the very similar extant species Caddo agilis : 1.64 mm for females, 1.9 mm for males (cf. Suzuki et al. 1977; Suzuki 1986). Minor differences in morphology can be observed in the Bitterfeld fossils, but compared to
extant taxa these are probably sexually dimorphic. For example, MB.A. 1655 ( Fig. 12 View Figures 12-14 ), without obvious spines or thorns on the pedipalp femur, is probably male, whereas MB.A. 1656 ( Fig. 14 View Figures 12-14 ), which has such spination, is probably female. The holotype of C. dentipalpus clearly has three femoral thorns ( Bishop and Crosby 1924: fig. 1). By comparison there are three such thorns in Caddo agilis females, whereas males have only one strong, finger-like protuberance (e.g. Gruber 1974: figs 18-22; Suzuki and Tsurusaki, 1983).
A distal mesal protuberance of the femur – as in MB.A. 1656 ( Fig 14 View Figures 12-14 : ap) – is of some significance. Visible in both Bishop and Crosby’s (1924: fig. 1) and Dunlop’s (2006: fig 2A) drawings of the C. dentipalpus holotype too, this apophysis has also been reported in C. agilis . In detail, Bishop (1949) reported this structure at the distal mesal angle of the pedipalp femur in juveniles from North America. Suzuki (1958) reported it from his (Japanese) females and Gruber (1974) figured it in adult females from North America. It appears to be generally absent from males ( Gruber 1974; Suzuki and Tsurusaki 1983). This femoral protuberance again emphasises the similarities – if not the conspecificity sensu Shear (1975) – between fossil and living Caddo harvestmen. The question of whether the amber examples merit a separate species is difficult to answer and further fossils would be welcome to test the stability of characters both between genders and instars. Bishop and Crosby’s (1924: fig. 1) illustrations also hint at a protuberance on the mesal side of the patella and a rather poorly defined feature in this area – or at least a group of setae – was observed here in MB.A. 1655 ( Fig. 12 View Figures 12-14 ). If this character could be confirmed in other specimens it would offer a possible diagnostic character compared to Recent species.
Conceivably, Eocene–Oligocene populations of European Caddo occupied specialized habitats and expressed subtle differences from modern Asian and North American forms which are difficult to assess in the available fossils. To complicate matters further, some modern Caddo species are thought to have arisen through neotony of isolated populations ( Shear 1975; Rambla 1980) and there are also frequent reports of parthenogenesis among the Recent fauna (e.g. Gruber 1974; Shear 1975; Suzuki 1976; Suzuki and Tsurusaki 1983) all of which may hinder the resolution of unequivocal apomorphies for the amber species. Shultz and Regier (2009) recently argued that C. agilis and its potential neonate C. peperella Shear, 1975 – both of which occur disjunctly in North America and Japan – evolved as distinct species, prior to their separation into American and Asian populations. Questioning the neotony hypothesis, they further discussed the potential role of paedo- and peramorphosis in understanding Caddo evolution and noted the need for further data from ancestral Caddo populations. With these provisions in mind, we prefer to retain C. dentipalpus as a separate taxon for the time being.
| MfN |
Museum für Naturkunde |
| MB |
Universidade de Lisboa, Museu Bocage |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.