
Capitella giardi ( Mesnil, 1897 )
|
publication ID |
https://doi.org/ 10.5281/zenodo.215310 |
|
publication LSID |
lsid:zoobank.org:pub:07C06068-9160-4AB4-AAF4-0451679D9F13 |
|
DOI |
https://doi.org/10.5281/zenodo.6175593 |
|
persistent identifier |
https://treatment.plazi.org/id/03913362-FF8C-FF8F-99BE-F93A990E48AF |
|
treatment provided by |
Plazi |
|
scientific name |
Capitella giardi ( Mesnil, 1897 ) |
| status |
|
Capitella giardi ( Mesnil, 1897) View in CoL
Figures 1 View FIGURE 1 A–C, 2 A–D, 3 C–E.
Capitellides giardi Mesnil, 1897: 441 View in CoL –443.
Capitella giardi View in CoL .- Warren 1976: 205 –206 (redescription).
Material examined. Oahu Island: Kahuku, aquaculture ponds, oyster farms, coll. R. Brock, Sept. 1988 (8, BPBM R3582); 15 Aug. 1988 (14, BPBM R3583); 25 Jan. 1989 (5). Hawaii Island: Kona coast with Oyster edulis and Boccardia proboscidea ( Bailey-Brock 2000) , with larvae, 15 Mar. 1990 (1, BPBM R3584); Anchialine Pond Preservation Survey, Jun. 2004, coll. R. Brock & A. Kam, Sta. 188R1 (18, BPBM R3585), Sta. MMIR 2 (33, BPBM R3586).
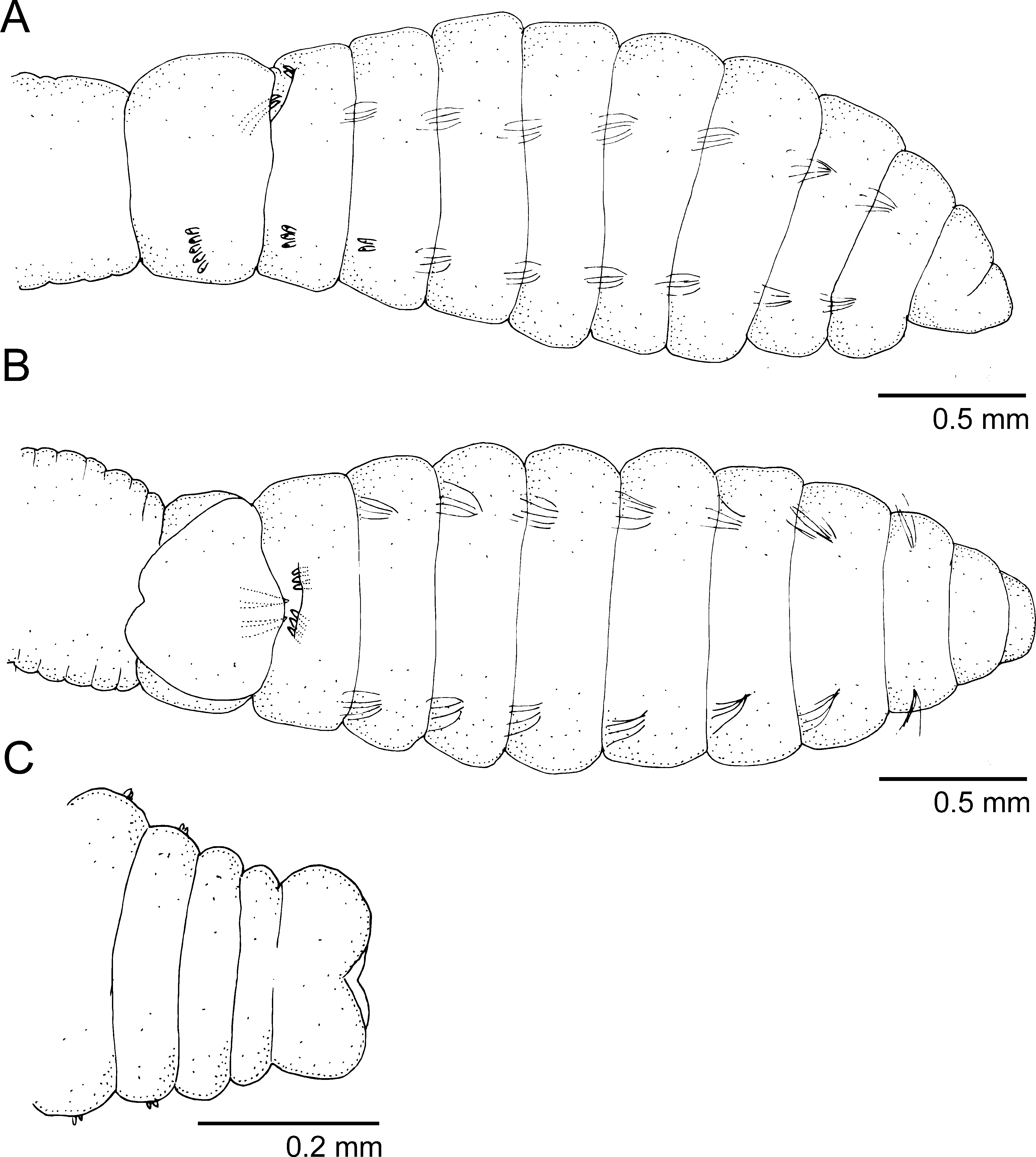
Description. Complete specimens 2.5–19 mm long, 0.1–0.8 mm wide for about 23–40 chaetigers. All specimens with external male characteristics (genital spines) but some bearing eggs in the coelom. Body elongate, wider on mid-thoracic chaetigers, gradually narrowing before pygidium; rounded dorsally and with ventral groove beginning after chaetiger 7; posterior end with neuropodia on shoulder-like projections. Color in alcohol white to pale yellow in most material but specimens collected from anchialine pools with brownish-orange speckles on anterior end, thoracic region and abdominal parapodial tori (see similar MGSP and Fig. 3 View FIGURE 3 C).
Prostomium conical, short, rounded anteriorly ( Figs 1 View FIGURE 1 A, 2B); nuchal organs dorso-laterally on prostomium, eyes not observed. Peristomium distinct dorsally from prostomium but not forming complete ring; dorsally as long as prostomium but slightly shorter than chaetiger 1 ( Figs 1 View FIGURE 1 A, B, 2B). Proboscis not observed.
Thorax with nine segments with variable arrangement of chaetae depending on growth stage. Four juveniles 2.5–4.0 mm long, 0.1–0.2 mm wide for 21–26 chaetigers. Thorax of juveniles with 1) chaetigers 1–3 with capillaries only and chaetigers 4–6 with mixed hooks and capillaries or 2) chaetigers 1–4 with capillaries only and chaetigers 5–6 with mixed hooks and capillaries. Developing embryos with up to 12 chaetigers and first three chaetigers with capillaries only. Mature individuals with chaetigers 1–6 with capillaries only, chaetiger 7 with notochaetae and neurohooks, chaetigers 8 and 9 with genital spines on notopodia and neurohooks ( Figs 1 View FIGURE 1 A, B, 2A). Thoracic segments uniannulate, similar in width from chaetiger 1–6, slightly wider on chaetiger 7, almost twice as wide on chaetigers 8–9; segments apparently distinct after chaetiger 5. Capillary chaetae numbering 4–5 per fascicle, unilimbate. Thoracic and abdominal hooded hooks similar; neuropodia of chaetiger 7–9 with 4–6 hooded hooks.
All specimens including juveniles, with genital spines on chaetigers 8–9 ( Figs 1 View FIGURE 1 A, B, 2A). Genital spines on mature individuals distinct; chaetiger 8 with two sets of 3–4 short spines facing posteriorly and chaetiger 9 with two sets of two robust spines facing anteriorly, spines sometimes superimposed; chaetiger 9 dorsally inflated ( Figs 1 View FIGURE 1 B, 2A).
Abdominal chaetae all hooded hooks; 3–4 notohooks and 5–6 neurohooks, similar in shape and length and reduced to 2–3 notohooks and 3–4 neurohooks posteriorly. Abdominal segments multi-annulated at least 3 wider than anterior thoracic segments. Noto- and neuropodia with well separated glandular tori pads; posterior neuropodial tori enlarged, glandular in mature specimens. Lateral organs and genital pores not observed. Hooded hooks with long hood, leaving only tip of main fang uncovered ( Fig. 2 View FIGURE 2 D). Hooks with moderate anterior shaft, slight node and multiple teeth; several rows of teeth visible in frontal view—three teeth basally, mid tooth enlarged, four in middle row and 3–4 in superior row ( Fig. 2 View FIGURE 2 D).
Pygidium with four inflated lobes and terminal anus ( Figs 1 View FIGURE 1 C, 2C, 3F).
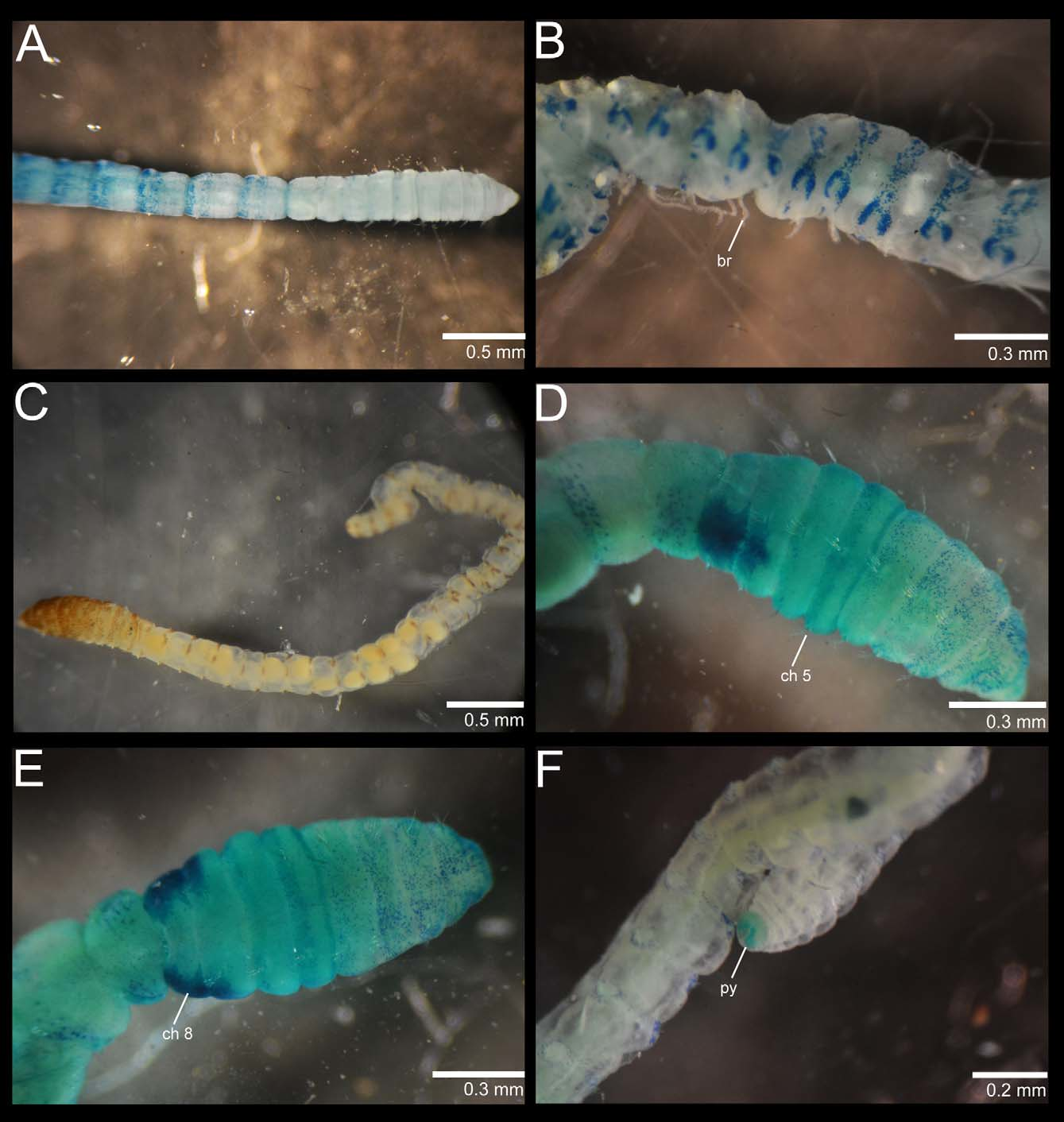
Methyl green staining pattern. All specimens (externally males) with similar staining reaction. Prostomium, peristomium and chaetigers 1–4 with dark speckles encircling segments but more abundant on edge of segments ( Fig. 3 View FIGURE 3 D, E); chaetigers 5–6 with a solid light green staining and chaetigers 7–8 with lateral glandular patches intensely stained ( Fig. 3 View FIGURE 3 D, E; genital duct? see Blake 2009). Chaetiger 9 and following abdominal segments with bands of speckles on posterior half of segment, encircling pre- and post-chaetal tori and forming complete rings ( Fig. 3 View FIGURE 3 D). Mid-ventral line on abdomen with sparse speckles and distinct ventral circles staining similar to parapodial tori. Pygidium darkly stained ( Fig. 3 View FIGURE 3 F). Juveniles stained similar to mature individuals except that lateral glandular patches were not visible.
Distribution. Capitella giardi is known from North Atlantic, Florida ( Warren 1976), Alaska ( Hartman 1961 as Capitella punctata ) and may have been introduced to Hawaii.
Remarks. This Capitella species was collected from oyster farms and may have been introduced to the Hawaiian Islands together with Polydora normalis (see Bailey-Brock 1990) and Boccardia proboscidea (see Bailey-Brock 2000). It agrees well with the available information of Capitella giardi in chaetal arrangement, coloration of some preserved specimens, and that both sexes have genital spines ( Warren 1976; Hartman 1961). The prostomium of the Hawaiian material is clearly distinct from the peristomium, which is elevated and incomplete ventrally. Warren (1976) points out the presence of a dorsal horizontal cleft for this species. In addition, chaetiger 7 has notochaetae and neurohooks in all mature specimens, distinguishing it from other closely related Capitella species. The MGSP is very distinct and may be unique for this species and its description herein should be compared to type material (if available) or material from the type locality, Wimereux, Pas-de-Calais, France ( Mesnil 1897).
Hermaphroditism is commonly found in populations of several species of Capitella and low population densities (as when a newly introduced species) may trigger hermaphroditic development of males ( Holbrook & Grassle 1984). This could be the reason why all specimens found had male characteristics.
Biology. Tubes had a fine layer of mucus to which fecal pellets adhere; frequently, two specimens were found inside each tube, with each one facing a separate entrance. Intracoelomic eggs were seen throughout but observed densely packed in chaetiger 9. Fertilized eggs were observed in the coelom and tubes. Embryos and larvae attach to the inner walls of tubes as reported in other Capitella species.

| BPBM |
Bishop Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Family |
|
|
Genus |
Capitella giardi ( Mesnil, 1897 )
| Magalhães, Wagner F. & Bailey-Brock, Julie H. 2012 |
Capitella giardi
| Warren 1976: 205 |
Capitellides giardi
| Mesnil 1897: 441 |