
Phyrella fragilis (Mitsukuri & Ohshima, 1912)
|
publication ID |
https://doi.org/10.11646/zootaxa.3760.2.1 |
|
publication LSID |
lsid:zoobank.org:pub:C0E4CA6E-6D93-477E-99F6-14FFB652A2A7 |
|
DOI |
https://doi.org/10.5281/zenodo.5041365 |
|
persistent identifier |
https://treatment.plazi.org/id/038F8787-C11B-435B-FF6C-EB34FAB8FAF6 |
|
treatment provided by |
Felipe |
|
scientific name |
Phyrella fragilis |
| status |
|
Phyrella fragilis View in CoL (Mitsukuri & Ohshima in Ohshima, 1912)
Figures 4–9 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 View FIGURE 8 View FIGURE 9 ; 27 a–c View FIGURE 27
Phyllophorus fragilis Mitsukuri & Ohshima View in CoL in Ohshima, 1912: 81–87. Pl. I, Fig. 3 View FIGURE 3 ; textfig 6. Syntypes UMUTZ-Ecn-H-Den- 144 (was 1658 in Mitsukuri & Ohshima, 9 specimens), Nishino-omote, Tanegashima, Stasuma; Satsukawa, Amami- Oshima (1 specimen); Sakibaru, near Naha, Okinawa island (12 specimens)
Phyrella fragilis: Heding & Panning, 1954: 185–186 View in CoL . Fig. 90 Thyonidiella oceana Heding & Panning, 1954:172–173 View in CoL , Fig. 82. Holotype: ZMUC HOL-253; type locality: Ambon, Moluccas, Indonesia
Thyonidiella oceana: Cherbonnier, 1988: 229–232 View in CoL , Fig. 102 A–H
Material examined: ZMUC HOL-253 (Holotype of Thyonidiella oceana ), Ambon, February 1922, coll. Th. Mortensen ; UF 4096 , Taiwan, Wanlitung , W of Kenting National Park, 0 m, 14 May 2005, coll. Kris Netchy & Robert Lasley ; UF 11011 , UF 11013 , UF 11014 , UF 11016 , Japan, Okinawa Island, Sunabe (adjacent to the type locality), 19 May 2011, coll. Yoshida Ryuta. WAM Z26012, Western Australia, Kimberley, Coulomb Point , - 17.3975, 122.148500, Tide pool, 8 October 2009, coll. A. Sampey GoogleMaps ; WAM Z26013 Western Australia, Kimberley, James Price Point , -17.501667, 122.143167, 7 October 2009, coll. G. Young. GoogleMaps
Description. External morphology. Body wall soft, fairly thin and covered with tube feet. Color in preservative, beige (UF 11011, UF 11013, UF 11014, UF 11016, ZMUC HOL-253, Fig. 5B View FIGURE 5 ) to gray (UF 4096) to orange (WAM Z26012, WAM Z26013), some with deep purple patches usually in the middle of the body and/or with small brown spots that can be fairly abundant all over body ( Fig. 5A View FIGURE 5 ); some specimens with darker coloration on both ends (e.g., UF 11011, Fig. 5A View FIGURE 5 ). Body arched, slightly U-shaped, generally cylindrical with a tapering posterior end. Largest animal (UF 4096) 65 mm along dorsum, 92 mm along venter, and 30 mm wide across midbody; most 40–55 mm along dorsum, 55–75 mm along venter, and 20+/- mm wide. Introvert retracted in all specimens examined, short (7 mm long, 4 mm wide for UF 4096), variable in color from almost white to dark brown. Tube feet same color as body wall and purple when originating in patches of this color; very abundant, evenly spread across radial and inter-radial areas, more abundant on venter and toward extremities; fairly large (~ 2 mm long, ~ 1 mm wide relaxed). 15–17 tentacles of varying size [UF 4096: 17 tentacles (10 in outer circle, 7 in inner circle), WAM Z26012: 16 tentacles (10 in outer circle, 6 in inner circle), WAM Z26013: 15 tentacles (9 in outer circle, 6 in inner circle), ZMUC HOL-253: 15 tentacles (disposition too difficult to discern)]. Cloacal membrane white. Cloaca surrounded by 5 small, lightly calcified anal teeth.
Internal anatomy. Polian vesicle single, white with sparse small brown spots; variable in size and with no clear relation to size of animal (4 mm in UF 4096,> 10 mm in all other dissected specimens). Stone canal inconspicuous, lightly calcified, at least proximally embedded in either the membrane surrounding the calcareous ring or the dorsal mesentery. Madreporite single, white, inconspicuous, small (<1 mm in diameter), spherical, either embedded in membrane surrounding calcareous ring, dorsal mesentery or free in the body cavity. Gonads well developed in mature individuals, in two tufts; composed of simple, up to 8 mm long, white or yellow tubes branching dichotomously up to a few times. Four of six dissected specimens examined were eviscerated, these retaining gonads, both respiratory trees, and a small anterior portion of the esophagus. Intestine long, distended when packed with fine, muddy sand. Respiratory trees white, extending almost entire body length.
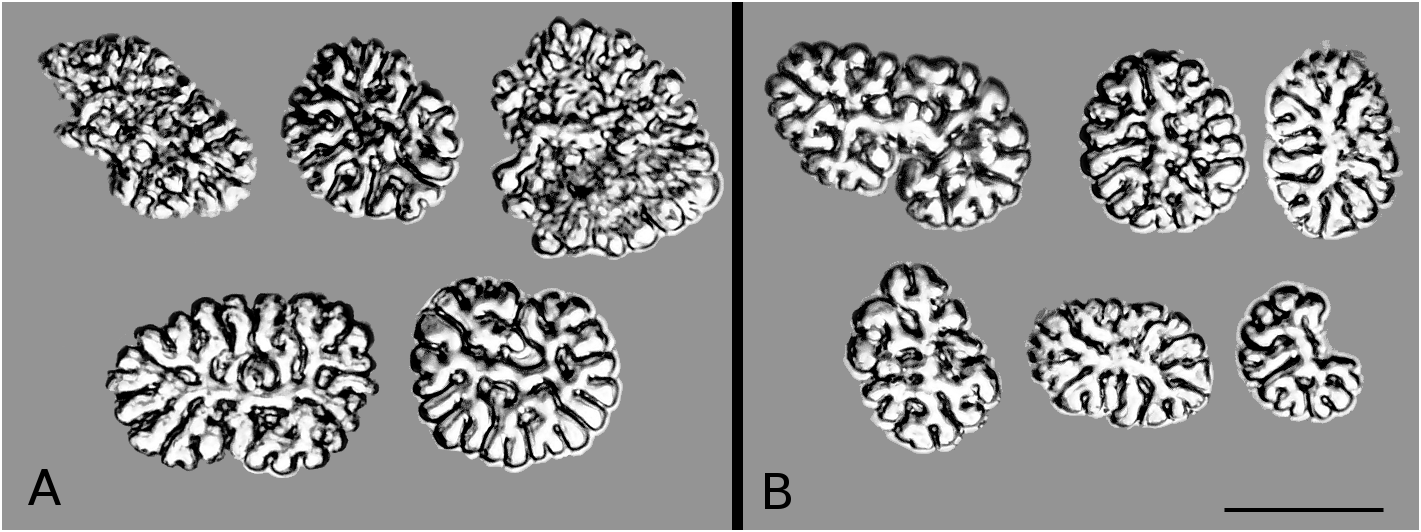
Calcareous ring. Calcareous ring embedded in a relatively thick membrane; radials and inter-radials typically highly fragmented elements; radial and inter-radial elements closely associated along most of their length ( Fig. 27a,b,c View FIGURE 27 ). Anterior margin of radial plates unequally divided by a well formed notch. Anterior margin of the inter-radial plates arrow-head shaped. Fragmentation of radial and inter-radial elements appears to increase with size; with inter-radials entire in one of the smallest specimens studied (WAM Z26012, Fig. 27b View FIGURE 27 ), to fragmented to 15–20 elements each, in largest (UF 4096, Fig. 27a View FIGURE 27 ). Radials extend posteriorly as long tails that tend to recurve at their tip.
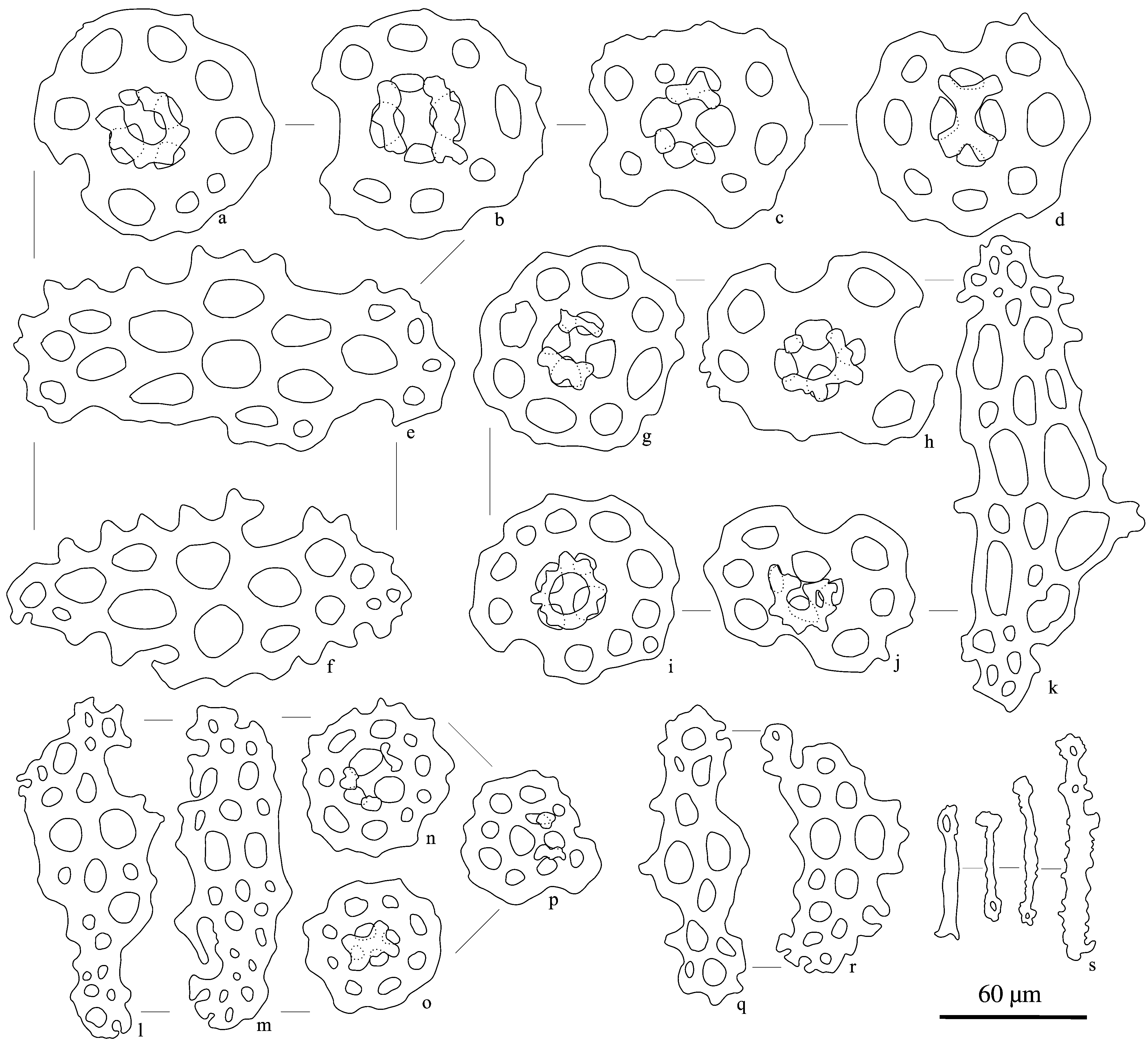
Ossicle assemblage. Dorsal and ventral body wall with tables only. Tables variable, disc with complete ( Fig. 7b View FIGURE 7 ) or incomplete rim ( Fig. 7f,d View FIGURE 7 ; Fig. 8a,d View FIGURE 8 ), 100–125 µ m in diameter; central perforation 10–35 µ m in diameter, surrounded by a ring of 8 perforations ( Fig. 7b,c View FIGURE 7 ; Fig. 8g View FIGURE 8 ), rarely with additional holes peripheral to these; margins slightly raised; spire formed by four pillars connected by a single cross-beam half way up, terminating in a narrow, often incomplete crown; irregular and incomplete tables also present, spire completely ( Fig. 7a,g View FIGURE 7 ) or partially wanting ( Fig. 8d, h View FIGURE 8 ), often with larger central perforation than regular tables. Podia with perforated plates 160–250 µ m long and 65–100 µ m wide, with holes decreasing in size toward periphery; margins serrated; serrations most developed along the more convex margin ( Fig. 7h–k View FIGURE 7 ; Fig. 8e,f,k View FIGURE 8 ). Cloacal body wall with tables and rosettes. Tables similar to those found elsewhere in body wall. Rosettes abundant, 20–30 µ m long ( Fig. 9A View FIGURE 9 ). Podia with shorter perforated plates, up to 150 µ m long ( Fig.7r View FIGURE 7 , Fig. 8q,r View FIGURE 8 ). Podia that flank anal teeth with rods, 40–65 µ m long ( Fig. 8s View FIGURE 8 ), and some rosettes. Introvert body wall with tables and rosettes (20–30 µ m in length) ( Fig. 9B View FIGURE 9 ); tables drop out toward tentacles. Tables similar to, but typically smaller (60–90 µ m) ( Fig. 7l–n View FIGURE 7 ; Fig. 8n–p View FIGURE 8 ) than those elsewhere in the body wall. Podia with perforated plates variable and mostly similar to those of body wall near cloaca ( Fig. 7o–q View FIGURE 7 ; Fig. 8l,m View FIGURE 8 ); and with rosettes, mostly toward the base of podia. Tentacles with rods, 40–95 µ m long, with no, one or a few perforations toward ends, some with serrations along margin ( Fig. 7s View FIGURE 7 ). No ossicles were observed in the cloacal, longitudinal and retractor muscles; intestine; cloacal wall; gonads; and respiratory tree.
Remarks. Mitsukuri noted in his original description (quoted in Ohshima, 1912) that the species has 15 tentacles (10 large and 5 small); however Ohshima adding to this description in the same paper notes that this species has “invariably” 20 tentacles. Heding & Panning (1954) followed Ohshima to assign this species to Phyrella based on having 20 tentacles. Specimens examined had 15–17 tentacles. This species can be distinguished from other Phyrella by the abundant, large tube feet, the regular tables in the body wall and the large serrations on the more convex margins of the perforated plates.
Synonymy of Thyonidiella oceana . Heding & Panning (1954) studied three specimens that they attributed to their Thyonidiella oceana , one from Ambon and two from Mauritius, designating the first as the holotype. The Mauritius specimens pertain to Phyrella ? ambigua (see below). Heding & Panning noted that the holotype is 25 mm long, appears contracted so that the body wall is thick, both mouth and anus are dorsal, and possesses 10 large and 5 small tentacles. They illustrated the calcareous ring showing that both radials and inter-radials are fragmented, and the inter-radials have posterior projections. They also noted that the body wall lacks ossicles, as they were able to recover only tube feet end plates. The absence of signs of corrosion on the latter suggested to the authors that the absence of ossicles was not an artifact. They end their description by providing notes from Mortensen who collected the Ambon specimen and indicated that it lives buried in the sand under rocks (presumably in the intertidal), and eviscerates readily.
Cherbonnier (1988) redescribed and illustrated the holotype. He noted that the specimens from Mauritius are devoid of ossicles except for tube feet end plates and rosettes in the cloacal region. This may explain why Heding & Panning did not find ossicles if they only examined the Mauritius specimens for ossicles. In his description, Cherbonnier confirmed Heding & Panning's description, and in particular the absence of ossicles in the ventral body wall of the holotype. However, he found and illustrated ossicles in the dorsal body wall. These include tables with a four pillar spire, and a crown which can be incomplete; perforated plates; and rosettes in the introvert and cloacal region. He concluded by noting that the original description is incomplete due to the lack of illustration of the dorsal body wall ossicles, and that the Ambon specimen appears to be Semperiella tenera ( Ludwig, 1875) . His concept of S. tenera was based on Heding & Panning account for this species, which in turn was not based on type material.
The ossicles and calcareous ring of the holotype of T. oceana , as illustrated by Cherbonnier (1988: Fig 102) are indistinguishable from Phyrella fragilis . Examination of the holotype of T. oceana ( Fig. 6 View FIGURE 6 ), confirms Cherbonnier's observations (absence of ossicles in the ventral body wall, tables and perforated plates in the dorsal body wall and tube feet, fragmented inter-radial elements in the calcareous ring), and we found no difference between it and specimens of P. fragilis taken near the type locality of the latter species.
Ecology & distribution. Phyrella fragilis is an intertidal species found under rocks in calcareous sand; the animal usually covers itself with large sand grains and pieces of shells. It is common in intertidal areas of Ilocos Norte province, Philippines (-17.501667, 122.143167), where it is commercially harvested and eaten by the local population who call it sorsorbot (R. Olavides, pers.comm.). Ohshima & Mitsukuri (1912) named the species for its propensity to eviscerate, noting: “As soon as the stone under which these animals live are lifted up, they throw off the viscera, even before they are touched in any way.” This species has the widest known distribution of any Phyrella , extending from Western Australia GoogleMaps , through Indonesia and the Philippines, to Taiwan and Okinawa.






No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Phyrella fragilis
| Michonneau, François & Paulay, Gustav 2014 |
Thyonidiella oceana: Cherbonnier, 1988: 229–232
| Cherbonnier, G. 1988: 232 |
Phyllophorus fragilis
| Ohshima, H. 1912: 81 |
Phyrella fragilis: Heding & Panning, 1954: 185–186
| Phyrella fragilis: Heding & Panning, 1954: 185–186 |
| Thyonidiella oceana Heding & Panning, 1954:172–173 |