
Phallocryptus tserensodnomi, Alonso, Miguel & Ventura, Marc, 2013
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3670.3.5 |
|
publication LSID |
lsid:zoobank.org:pub:1773E3DE-6C8D-42C2-85AA-54A45D791801 |
|
DOI |
https://doi.org/10.5281/zenodo.6153021 |
|
persistent identifier |
https://treatment.plazi.org/id/038E87CC-FF8A-BA73-5CBB-4ACDFBF66595 |
|
treatment provided by |
Plazi |
|
scientific name |
Phallocryptus tserensodnomi |
| status |
sp. nov. |
Phallocryptus tserensodnomi View in CoL sp. nov.
( Figs 2–5 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 )
Etymology. Species named after Jimee Tserensodnom in recognition of his contributions to the knowledge of the Mongolian lakes, particularly the production of the first catalogue including more than 3,000 lakes ( Tserensodnom 2000).
Type locality. Shorvog nuur, code 342AR, Arkhangai aimag, Mongolia (47°39’0.54”N; 102°22’11.0”E).
Type material. Holotype. Undissected mature male preserved in 4% formaldehyde vial ( MNCN 20.02/ 17242), coll. M. Alonso, September 2009.
Allotype. Undissected mature female preserved in 4% formaldehyde vial ( MNCN 20.02/17243), coll. M. Alonso, September 2009.
Paratypes. 6 males and 10 females preserved in 4% formaldehyde vial ( MNCN 20.02/17244), coll. M. Alonso, September 2009.
Comparative material examined. All in MA collection. Uizen nuur, code: 322SU, Sukhbaatar aimag, Mongolia (45 ° 21’37.5”N; 113 ° 18’16.2”E), September 2006; Mendbayar nuur, code 1874 DOD, Dornod aimag, Mongolia (47 ° 51’51.7”N; 117 ° 57’58,6”), September 2006; Khalkh Gol bagiin nuur 2, code 1875 DOD, Dornod aimag, Mongolia (47 ° 49’22.2”N; 117 ° 54’40.0”E), September 2006; Shiliin nuur, code 142TU, Tuv aimag, Mongolia (47 ° 01’18.8”N; 106 ° 07’31.7”E), September 2009; Khar nuuriin toirom, code 11TU, Tuv aimag, Mongolia (48,21271 ° N; 104,90981 ° E), August 2012. Information and pictures of sampling sites can be easily accessed on the web (at http://www.geodata.es/mongolian_lakes/).
Diagnosis. Male. Frontal appendages short, provided with small conical outgrowths. Distal antennameres long and narrow, evenly curved, sickle-shaped, reaching to IV–V thoracopods IV–V. Distal fleshy process on labrum evenly curved forwards. Short stout spine-like projections present at each side of basal portion of gonopods. Distal margin of abdominal segments each provided with pair of large ventrolateral spine-like projections. Abdominal somites each with pair of medial spine-like projections on posterior margin; projections on first segment large, remainder smaller and progressively reduced towards posterior.
Female. Second antennae lamellar, broad, three times longer than wide, with broad pointed apex. Brood pouch fusiform, not protruding laterally from genital segments. Eggs sub-spherical with surface sculptured with shallow angular elongated depressed facets limited by blunt ridges.
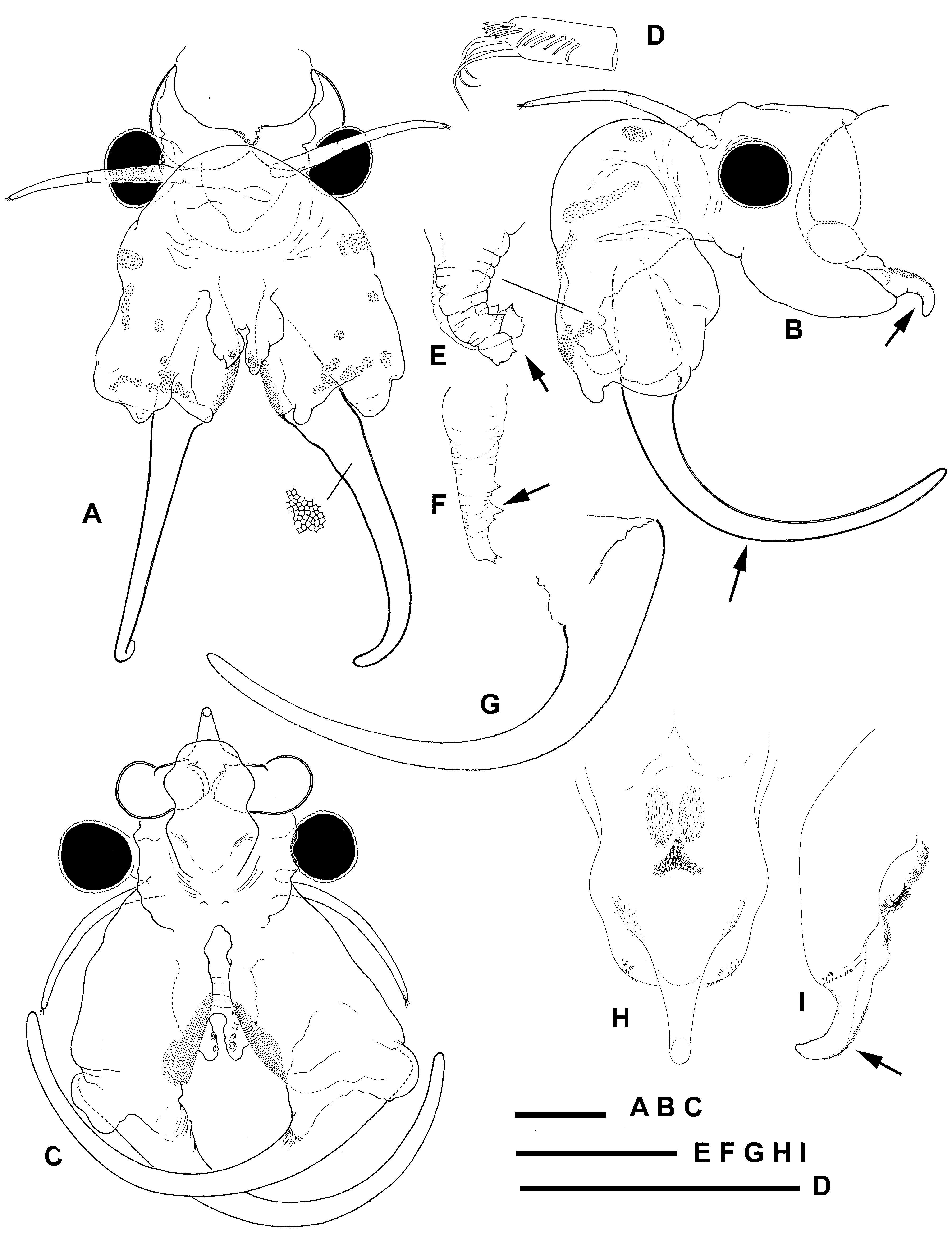
Description. Male. Body unpigmented. Head ( Fig. 2 View FIGURE 2 A–C) with rounded anterolateral angle barely covering eyestalk. Nuchal organ apparently absent. Eyes spherical with diameter longer than corresponding eyestalk.
First antennae ( Fig. 2 View FIGURE 2 A, D) filiform, three times longer than eye diameter and shorter than proximal antennamere of second antenna. Distal end with three subdistal setae similar in length, each twice as long as broad. Longitudinal comb of eleven short aesthetascs disposed distally on lateral margin of antenna as figured.
Second antennae ( Fig. 2 View FIGURE 2 A–C) long, capable of extending backwards to thoracopod IV or V. Proximal antennamere soft and corrugated, with patches of microtuberculate integument and sensillae distributed on dorsal and mediodistal surface as figured. Distal antennamere narrow, twice length of proximal counterpart, evenly curved backwards, sickle-shaped and tapering into blunt apex. Frontal appendage short, half length of proximal antennamere, provided with small ventral conical outgrowths ( Fig. 2 View FIGURE 2 E, F).
Labrum ( Fig. 2 View FIGURE 2 B, H, I) subtrapezoidal, lacking distal protuberances; terminal fleshy process elongate and evenly curved forwards, tapering distally. Triangular setulose pad placed midway on ventral surface.
Phyllopodia with gross structure typical for genus ( Fig. 3 View FIGURE 3 A–G). First thoracopod ( Fig. 3 View FIGURE 3 A) attaining two-thirds size of fifth thoracopod ( Fig. 3 View FIGURE 3 E); eleventh thoracopod ( Fig. 3 View FIGURE 3 F) 0.6 times as large as fifth thoracopod; rest of thoracopods subsimilar in size. Praepipodite (PE) oval, entire, with small notch placed midway of outer margin. Epipodite (EP) oval with smooth margin. Exopodite (EX) broad, “D”-shaped with plumose marginal setae. Endopodite (EN) broad; inner margin straight with marginal setae simple and sclerotized; length of setae variable both among individuals and between thoracopods of same specimen (even between left and right counterparts of same pair). First endite of thoracopods I–X with two submarginal spine-like setae on anterior surface, distal one with tiny spine-like seta at base ( Fig. 3 View FIGURE 3 D); counterparts on endite of eleventh thoracopod reduced. Second endite with two proximal setae submarginally on anterior surface, one of them reduced. Third and fourth endites of all limbs ( Fig. 3 View FIGURE 3 G; I–XI) each with two submarginal spine-like setae on anterior surface, one of which reduced, and with three and two long plumose setae on posterior surface, respectively. Fifth endite with one submarginal spinelike seta on anterior surface, and two long plumose setae on posterior surface.
Genital segments ( Fig. 4 View FIGURE 4 A, B, D) only slightly expanded and partially fused. Ventral surface of first segment smooth. Second genital segment ( Fig. 4 View FIGURE 4 D) with short stout acute spine-like projection similar to those on abdominal segments placed posterolaterally at each side on ventral surface. Basal portion of gonopods short and rigid, microtuberculate, with inflated, rounded medial surface covered with densely set short chitinized denticles. Eversible part of gonopods consisting of unique short boom-like process provided with longitudinal row of denticles proximally on medial margin plus cluster of shorter denticles subdistally at each side ( Fig. 4 View FIGURE 4 D, E).
Abdominal segments ( Fig. 4 View FIGURE 4 A, B) bearing warty outgrowths provided with sensillae distributed as follows: posterodorsal pair on the first, third and fifth segments; posterolateral pair on the second and fourth segments; and two posterodorsal and one posterolateral on the sixth segment. In addition, all segments with pair of ventrolateral curved spine-like projections at each side close to posterior margin, sometimes appearing duplicate in most posterior segments ( Fig. 4 View FIGURE 4 C). First segment with pair of large mid-ventral straight spine-like projections close to posterior margin; other segments with similar but reduced projections, although sixth frequently unarmed.
Cercopods five times longer than broad and three times longer than telson. Terminal setae implanted as in Fig. 4 View FIGURE 4 F.
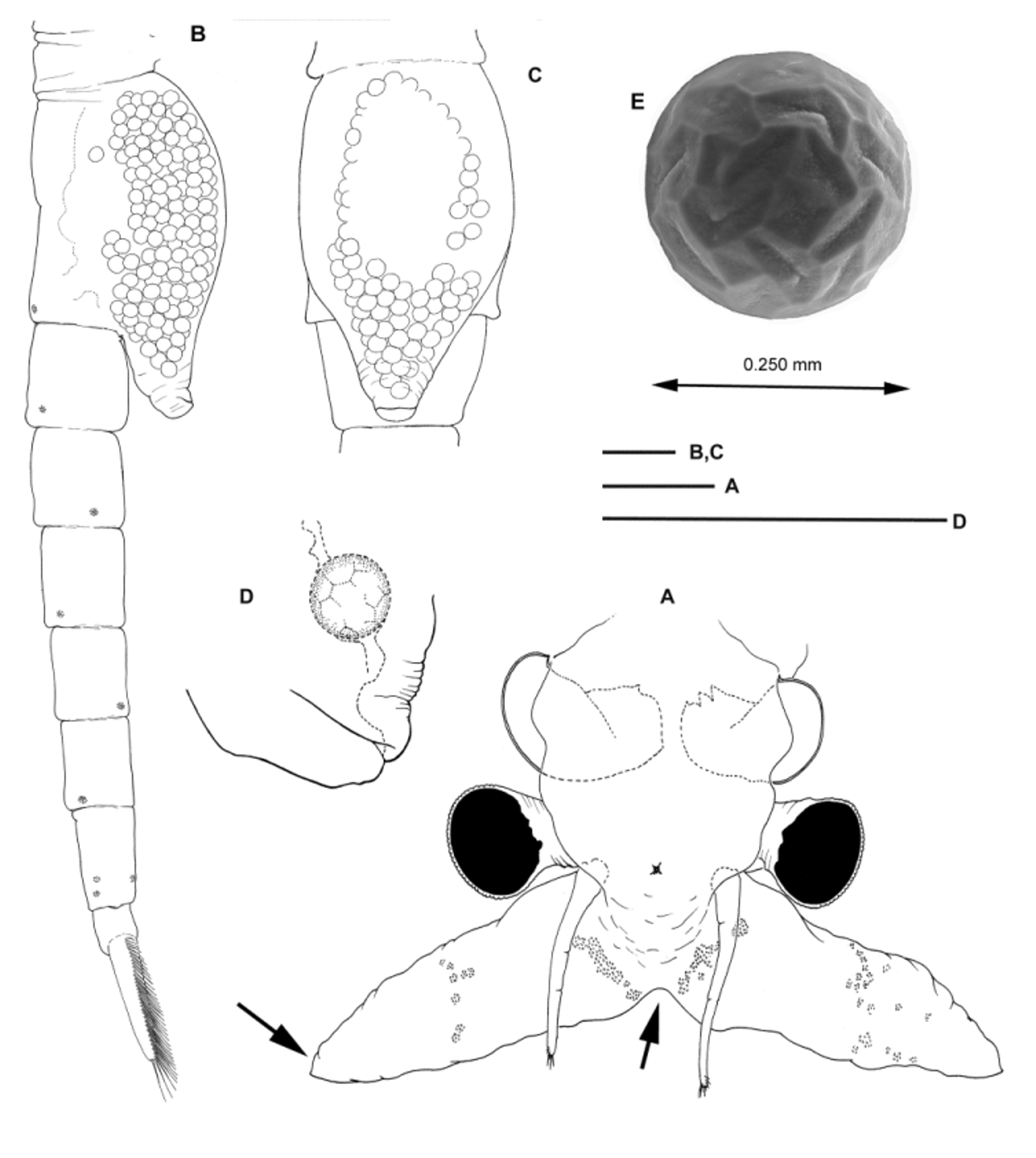
Female. First antennae approximately twice as long as eye diameter and twice shorter than second antennae ( Fig 5 View FIGURE 5 A).
Second antennae ( Fig. 5 View FIGURE 5 A) soft, lamellar, three times longer than wide, with broad pointed apex. Patches of microtuberculate integument sparsely set on surface of antenna as figured. Counterparts fused medially at base, and forming obtuse angle in frontal view.
Genital segments ( Fig. 5 View FIGURE 5 B, C) completely fused. Brood pouch fusiform, not protruding laterally, tapering posteriorly; apex reaching midway of second abdominal segment. Gonopore as illustrated in Fig. 5 View FIGURE 5 D, produced postero-ventrally.
Abdominal segments unarmed, lacking spine-like processes ( Fig. 5 View FIGURE 5 B).
Eggs. Numerous, more than 500 per brood pouch. Diameter 250 μm. Sub-spherical, with surface sculptured with shallow angular elongated depressed facets limited by blunt ridges ( Fig. 5 View FIGURE 5 E).
Size. Total length of holotype (including cercopods setae) 24.58 mm (head plus thorax 10.80 mm, abdomen 13.78 mm). Length of allotype 25.58 mm. Largest specimen recorded (female) 33.65 mm.
Differential diagnosis. Among members of the genus Phallocryptus , P. spinosa is closest to the new species. Nevertheless, P. tserensodnomi sp. nov. can be easily distinguished based on the following features: (1) frontal appendage is provided with small ventral conical outgrowths, unlike P. spinosa , which displays smooth frontal appendages; (2) second antennamere is evenly curved, sickle-shaped, more similar to that of P. wrighii and differing from the condition in P. spinosa , where the main curvature of segment is attained at its proximal half; (3) the distal fleshy process on the labrum evenly curved forwards and taper distally versus process “L” shaped in P. spinosa ; (4) short stout spine-like projections are present on each side of basal portion of gonopods (versus projections absent in P. spinosa ); (5) the first four abdominal segments have a distinctive midventral bulge on the integument on the anterior margin in P. s p i n o s a (versus bulges absent in the new species); (6) the female second antennae are shorter and wider than in P. spinosa , and without the acute recurved antenna tip characteristic of this species. Genetic differences between both taxa are described in the following section.
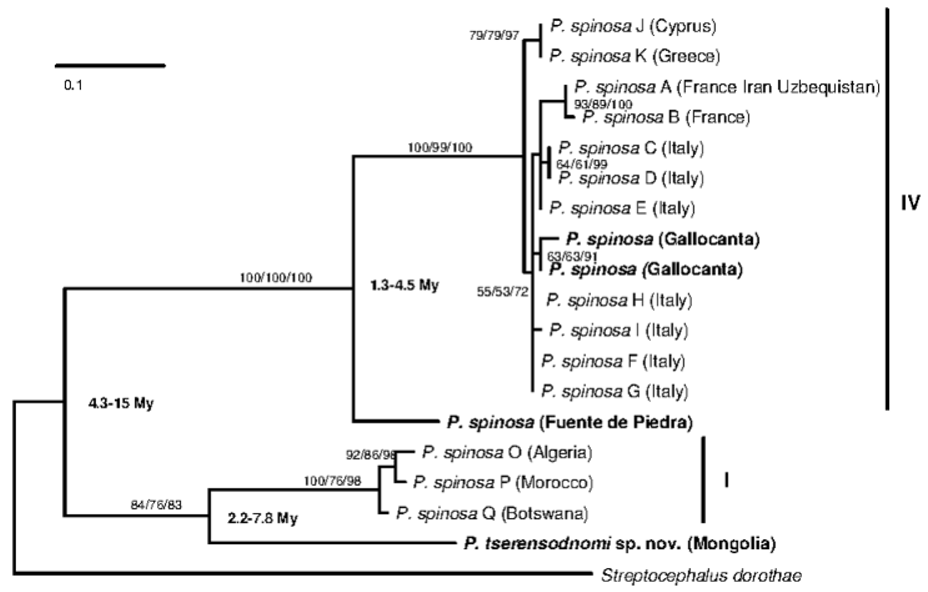
Genetic analyses. The COI mitochondrial gene of P. tserensodnomi sp. nov. clearly showed a considerable divergence from all other so-far genetically characterised species/populations of the genus. Phallocryptus tserensodnomi sp. nov. is divergent from the genetically most similar species, P. s p i n o s a, by 20% from the group IV; 19.2% from the Fuente de Piedra population; and by 10.9% from the group I (all Kimura 2-parameter distances) ( Fig. 6 View FIGURE 6 ). Group I was actually closer to P. tserensodnomi sp. nov. than to the group IV (19.2 %) or to the Fuente de Piedra population (18.2 %). Among the two populations analysed in this study (the Iberian sites of Gallocanta and Fuente de Piedra), the Gallocanta population was very similar to haplotypes belonging to the clade IV of Ketmaier et al. (2008), with 0.6 % divergence, while the population of Fuente de Piedra was more differentiated (6.7 %).
No variation in sequences of COI was observed in the two analysed individuals of P. tserensodnomi sp. nov. The GTR+G model of nucleotide substitution consistently performed best among the different approaches to model selection, based on the 508 bp alignment of COI sequences. All applied methods of phylogenetic reconstruction supported the sister relationship between the new species and P. spinosa despite their relatively high divergence. In addition, it suggests that the group I as described by Ketmaier et al. (2008) has a monophyletic origin with P. tserensodnomi sp. nov. Tentative molecular clock divergence time suggests that the separation between P. tserensodnomi and P. spinosa occurred between 4 and 15 Mya.
Distribution and ecology. So far P. tserensodnomi sp. nov. is a Mongolian endemic. It has been found in six lakes on the eastern half of the country, in the steppes and dry steppes (annual rainfall less than 350 mm) of the provinces of Arkhangay, Tuv, Sukhbaatar and Dornod, at altitudes from 588 to 1,346 m a.s.l. It is remarkable that the species has not been recorded in the western half of the country despite extensive sampling and presence of many suitable habitats, where its most common associated species are found. The lakes that it inhabits are shallow, temporary or permanent, but the latter freeze totally in the extremely cold Mongolian winter and therefore behave also as temporary. Waters are athalassic mesosaline, with salinities ranging between 10.8 and 25 g /L. Dominant ions are carbonate and/or chloride, and sodium. Water turbidity is high due to presence of suspended clay particles that are generally gray-coloured. No submerged macrophytes were recorded in any of these lakes. The accompanying crustacean community was composed of characteristic halophile species in Mongolia ( Alonso 2010): the fairy shrimps Artemia cf. sinica Cai, 1989 , Galaziella mongoliana (Uéno, 1940) , Branchinecta orientalis Sars, 1901 , and Branchinectella media Schmankewitsch, 1873 ; the cladocerans Daphnia triquetra Sars, 1903 , D. similis Claus, 1876 , Moina salina Daday, 1888 , M. brachiata (Jurine, 1820) and Macrothrix gr. hirsuticonis Norman & Brady, 1867; the copepods Metadiaptomus asiaticus (Ul’yanin, 1875) and Thermocyclops kawamurai Kikuchi, 1940 ; and some unidentified ostracods. It is remarkable that P. tserensodnomi sp. nov. normally co-occurs with some of the aforementioned fairy shrimps, in one case with three of them.


| MNCN |
Museo Nacional de Ciencias Naturales |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |