
Halopteris campanula ( Busk, 1852 )
|
publication ID |
https://doi.org/ 10.5252/z2009n1a3 |
|
persistent identifier |
https://treatment.plazi.org/id/038E87B3-9651-FFEB-ECCD-FC40AC764A18 |
|
treatment provided by |
Marcus |
|
scientific name |
Halopteris campanula ( Busk, 1852 ) |
| status |
|
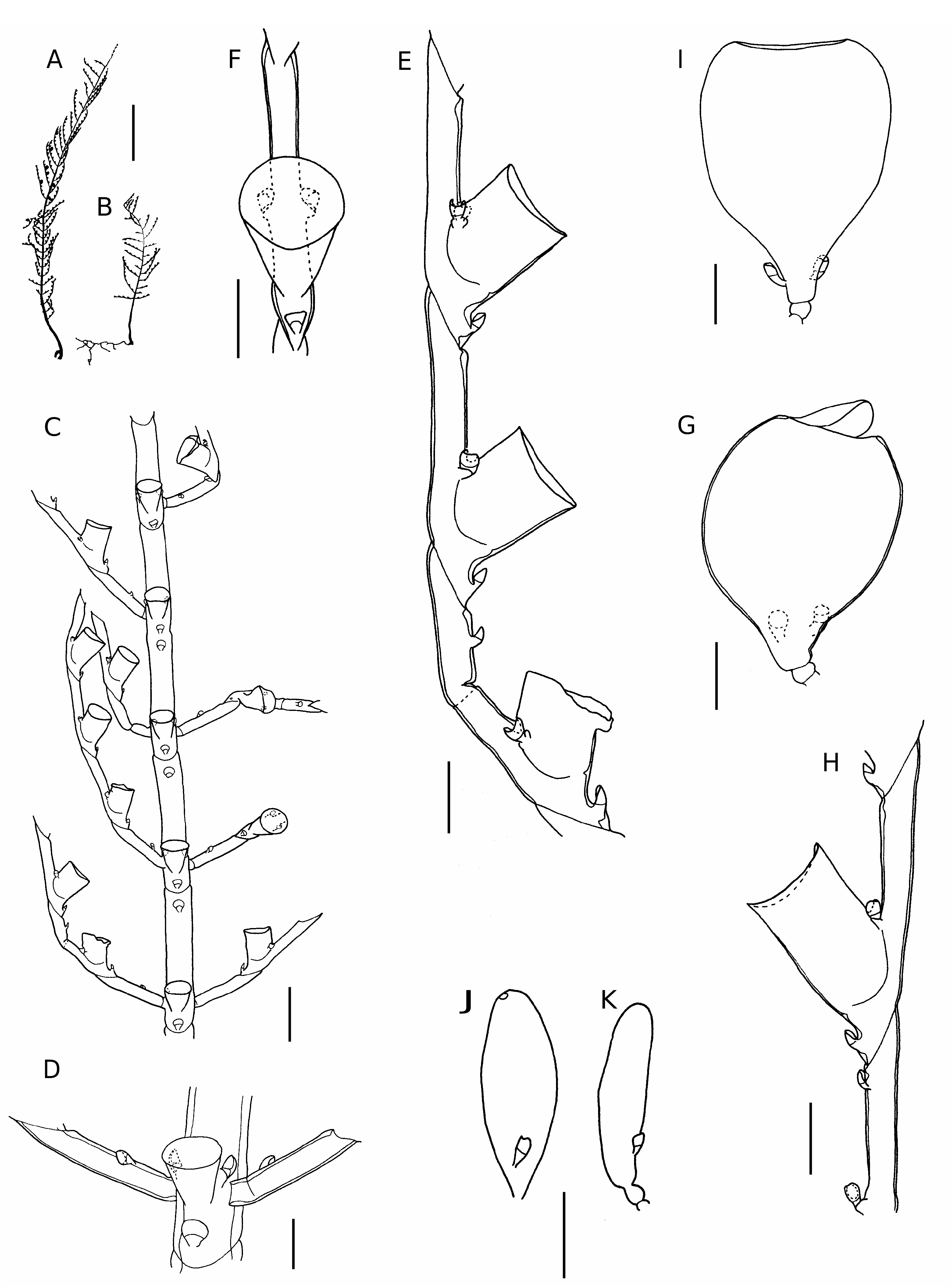
Halopteris campanula ( Busk, 1852) ( Figs 5 View FIG ; 6 View FIG ; Table 6)
Plumularia campanula Busk, 1852:401 .— Bale 1884: 124, pl. 10, fig. 5; 1888: 776, pl. 20, figs 1-6. — Billard 1913: 17, pl. 1, figs 11-13. — Hirohito 1974: 33, fig. 15.
Plumularia indivisa Bale, 1882: 39 , pl. 15, fig. 1.
Plumularia laxa Allman, 1883: 19 , pl. 1, figs 5, 6.
Plumularia rubra von Lendenfeld, 1885: 476 , pl. 13, figs 11, 12, pl. 14, fig. 15. — Bale 1888: 778, pl. 20, figs 1-6.
Plumularia torresia von Lendenfeld, 1885: 477 , pl. 13, figs 13, 14, pl. 14, fig. 16.
Plumularia zygocladia Bale, 1914: 171 , pl. 24, fig. 2.
Plumularia campanula var. geelongensis Mulder & Trebilcock, 1916: 76 , pl. 11, figs 2-2c.
Halopteris campanula var. zelandica Totton, 1930: 219 , text-fig. 57.
Halopteris campanula – Leloup 1938: 20, fig. 14. — Rees & Vervoort 1987: 124, fig. 26. — Schuchert 1997: 99, fig. 35. — Vervoort & Watson 2003: 353, figs 85G, 86A-E.
Halopteris campanula var. campanula Ralph, 1961: 47 .
Halopteris zygocladia – Schuchert 1997: 119, fig. 43.
MATERIAL EXAMINED. — New Caledonia. LAGON, stn 0420, 22°44.2’S, 167°08.9’E, 345 m, 24.I.1985, 25 stems rising from stolon, 3 with a few gonothecae ( MNHN).
BIOCAL 1, stn DW 77, 22°15.329’- 22°15.659’S, 167°15.405’- 167°15.498’E, 440 m, 5.IX.1985, 2 colonies on stem of Lytocarpia myriophyllum (Linnaeus, 1758) , no gonothecae ( MNHN). — Stn CP 108, 22°02.522’- 22°02.735’S, 167°05.680’- 167°06.050’E, 335 m, 9.IX.1985, many stems, 1 with damaged female gonothecae ( MNHN). — Stn CP 110, 22°12.383’- 22°13.315’S, 167°06.434’- 167°09.936’E, 275-320 m, 9.IX.1985, colonies on worm tube and stem of hydroid, 1 colony with female gonothecae ( MNHN).
MUSORSTOM 4, stn DW 162, 18°35.0’S, 163°10.3’E, 525 m, 16.IX.1985, several stems on gorgonid axis, some with male and female gonothecae (MNHN). — Stn CP 171, 18°57.8’S, 163°14.0’E, 435 m, 17.IX.1985, 10 colonies on various objects and other hydroids, some damaged gonothecae (MNHN).— Stn CP 180, 18°56.8’S, 163°17.7’E, 450 m, 18.IX.1985, many colonies attached to stems of other hydroids, on sponge and also some detached stems, with 6 female gonothecae (MNHN).
LAGON,stn 0599, 22°17.0’S, 167°06’E, 50 m, 5.VIII.1986, single stem with damaged male gonothecae (MNHN).
CHALCAL 2, stn DW 76, 23°40.50’S, 167°45.20’E, 470 m, 30.X.1986, 1 colony with several stems on sponge, with male and female gonothecae (MNHN).
CALSUB, stn PL 19, 22°46’S, 167°20’E, 416- 404 m, 10.III.1989, 4 stems, no gonothecae (MNHN).
Loyalty Islands. MUSORSTOM 6, stn DW 391, 20°47.35’S, 167°05.70’E, 390 m, 13.II.1989, 2 colonies with several stems, no gonothecae (MNHN). — Stn DW 461, 21°06.00’S, 167°26.20’E, 240 m, 21.II.1989, 1 stem without gonothecae (MNHN).
Norfolk Ridge. SMIB 4, stn DW 51, 23°41.3’- 23°40.5’S, 168°00.6’- 168°00.7’E, 245-260 m, 9.III.1989, 2 mutilated stems on alcyonarian axis, no gonothecae (MNHN). — Stn DW 53, 23°40.1’- 23°39.5’S, 167°59.9’- 168°00.3’E, 250-270 m, 9.III.1989, 1 colony with female gonothecae (MNHN). — Stn DW 55, 23°21.4’- 23°21.4’S, 168°04.5’- 168°04.8’E, 215-260 m, 9.III.1989, several colonies and fragments on sponge and Bryozoa, without gonothecae (MNHN). — Stn DW 56, 23°20.6’- 23°21.1’S, 168°05.2’- 168°04.7’E, 230-260 m, 9.III.1989, 1 colony with several stems, with female gonothecae (MNHN). — Stn DW 57, 23°21.5’- 23°21.0’S, 168°04.6’- 168°04.5’E, 210-260 m, 9.III.1989, 3 stems without gonothecae (MNHN). — Stn DW 64, 22°55.3’- 22°54.9’S, 167°16.4’- 167°15.5’E, 455- 460 m, 10.III.1989, many colonies without gonothecae (MNHN).
SMIB 5, stn DW 72, 23°42.0’S, 168°00.8’E, 400 m, 7.IX.1989, 1 mutilated stem without gonothecae (MNHN).— Stn DW 76, 23°41.2’S, 168°00.5’E, 280 m, 7.IX.1989, small fragment, no gonothecae (MNHN). — Stn DW 78, 23°40.8’S, 168°00.2’E, 245 m, 7.IX.1989, 2 colonies, no gonothecae (MNHN). — Stn DW 85, 22°20.0’S, 169°42.9’E, 260 m, 13.IX.1989, small fragment with a male gonotheca (MNHN). — Stn DW 93, 22°20.0’S, 168°42.3’E, 255 m, 13.IX.1989, 1 colony without gonothecae (MNHN).— Stn DW 95, 22°59.7’S, 168°19.8’E, 200 m, 14.IX.1989, 2 stems without gonothecae (MNHN). — Stn DW 101, 23°21.2’S, 168°04.9’E, 270 m, 14.IX.1989, many colonies attached to sponge and some fragments (some damaged), with female gonothecae (MNHN).— Stn DW 102, 23°19.6’S, 168°04.7’E, 305 m, 14.IX.1989, 2 fragments without gonothecae (MNHN).
DISTRIBUTION. — Rees & Vervoort (1987) consider Halopteris campanula to be restricted to tropical and subtropical parts of the Indo-west Pacific.
This species has been recorded from Japan ( Leloup 1938; Hirohito 1974), New Zealand ( Totton 1930 as H. campanula var zelandica ; Schuchert 1997; Vervoort & Watson 2003), Australia ( Busk 1852; Bale 1884, 1914 as H. zygocladia ), Malay Archipelago ( Billard 1913), Gulf of Aden ( Rees & Vervoort 1987) and Gulf of Suez and Red Sea ( Billard 1933).
The present material comes from New Caledonia, Norfolk Ridge and Loyalty Islands, and was collected between 50 and 525 m depth.
DESCRIPTION (COLONIES FROM SMIB 5,
STN DW 101)
Colonies formed by plumose cormoids arising from a tubular hydrorhiza.Stem monosiphonic, basal part with some ahydrothecate internodes separated by transverse nodes; the last ahydrothecate internode separated from rest of stem by an oblique node and provided with some frontal nematothecae. Remain- der of stem composed of hydrothecate internodes separated by oblique nodes. Each segment with one hydrotheca and four nematothecae: one mesial inferior, a pair of laterals and one mesial superior.
Hydrocladia arise laterally of hydrothecae of stem internodes on short, often indistinct apophyses. Hydrocladial arrangement is opposite basally and alternating right and left in middle and distal parts of stem. First internode of hydrocladium with oblique distal node and one or two nematothecae. Rest of hydrocladium composed of hydrothecate internodes separated by oblique nodes. Each segment with one hydrotheca and four nematothecae with the same distribution as on the stem.
Hydrotheca of stem and hydrocladium identical, almost cylindrical, walls slightly narrowing towards hydrothecal base; abcauline wall straight; adcauline wall adnate for half its length, rim circular, sometimes slightly flaring.
Mesial inferior nematotheca not reaching hydrothecal base and with adcauline wall of upper chamber missing. Lateral nematothecae short, bithalamic, placed on small apophyses, apical chamber globular and with deeply scooped adcauline wall, basal chamber small. Other nematothecae on stem and hydrocladia smaller and similar to mesial inferior nematothecae. Lateral nematothecae of stem with basal chamber better developed than those of hydrocladia.
Male and female gonothecae arising on same cormoid, inserted on internode next to hydrothecal base. Female gonothecae laterally compressed, pear-shaped, with oval terminal operculum; basal part narrowing to a pedicel composed of two short internodes, with two nematothecae of which adcauline wall of upper chamber is deeply scooped. Male gonothecae smaller than female, oblong, with small circular terminal aperture; basal part with one nematotheca placed on short pedicel similar to female gonotheca.
VARIABILITY
In youngest parts of some hydrocladia the nodes between hydrothecate internodes are only indistinctly visible.In apical parts of the colonies the supracalycine nematotheca is frequently absent in many axial and hydrocladial internodes, but this nematotheca never lacks in the basal parts. Many colonies sometimes have one supracalycine nematotheca in apical part, as the results of damage and posterior regeneration; nevertheless, the material from SMIB 4, stn DW 56 and SMIB 5, stn DW 85 has one supracalycine nematotheca in some internodes, but without signs of damage.
The material of SMIB 4, stn DW 57 is composed of three axis, one with supracalycine nematothecae but in the others the presence of this nematotheca is quite rare.
The number of supracalycine nematothecae in axial internodes varies between 0 and 2.
REMARKS
The material studied is characterized by the arrangement of the hydrocladia, opposite in basal parts of the colony and alternate in middle and distal parts. Besides, in the majority of the material the supracalycine nematotheca is absent; nevertheless, the morphology of colony and gonothecae, when present, is identical with material with supracalycine nematothecae.
Schuchert (1997) indicated that hydrothecae, nematothecae and female gonothecae of Halopteris campanula ( Busk, 1852) are indistinguishable from those of H. zygocladia ( Bale, 1914) , and the only difference between both species is the arrangement of hydrocladia, alternate in H. campanula and opposite in H. zygocladia . The author considers H. campanula a variable species but keeps both species apart because in his opinion the arrangement of hydrocladia, opposite or alternate, is a reliable character to separate species.
Our material shows the characteristic features of H. campanula , but with opposite hydrocladia in basal parts and alternate in middle and distal parts of the colony. The presence of both types of hydrocladial arrangements in the same colonies demonstrates that this character is variable too and therefore unsuitable for specific differentiation. In consequence we propose the inclusion of H. zygocladia in the synonymy of H. campanula .
In the absence of supracalycine nematothecae our material approaches H. gemellipara Millard, 1962 , but as all morphological characters of trophosome and gonosome agree with those of H. campanula it has been included in that species. Besides, lateral nematothecae of axis and hydrocladium are always two-chambered ( Figs 5 View FIG ; 6 View FIG ), despite the fact that their basal chamber can be strongly reduced; in H. gemellipara the lateral nematothecae are one-chambered. Development of the basal chamber of lateral nematothecae in H. campanula is variable ( Schuchert 1997), it may almost completely disappear. In our opinion, H. gemellipara is close to H. campanula , but as the gonosome of H. gemellipara is unknown it is necessary to study new material before drawing final conclusions.

| MNHN |
Museum National d'Histoire Naturelle |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Halopteris campanula ( Busk, 1852 )
| Agís, José Ansín, Vervoort, Willem & Ramil, Fran 2009 |
Halopteris zygocladia
| SCHUCHERT P. 1997: 119 |
Halopteris campanula var. campanula
| RALPH P. M. 1961: 47 |
Halopteris campanula
| VERVOORT W. & WATSON J. E. 2003: 353 |
| SCHUCHERT P. 1997: 99 |
| REES W. J. & VERVOORT W. 1987: 124 |
| LELOUP E. 1938: 20 |
Halopteris campanula var. zelandica
| TOTTON A. K. 1930: 219 |
Plumularia campanula var. geelongensis
| MULDER J. F. & TREBILCOCK R. E. 1916: 76 |
Plumularia zygocladia
| BALE W. M. 1914: 171 |
Plumularia rubra von Lendenfeld, 1885: 476
| BALE W. M. 1888: 778 |
| LENDENFELD R. & VON 1885: 476 |
Plumularia torresia von Lendenfeld, 1885: 477
| LENDENFELD R. & VON 1885: 477 |
Plumularia laxa
| ALLMAN G. J. 1883: 19 |
Plumularia indivisa
| BALE W. M. 1882: 39 |
Plumularia campanula
| HIROHITO 1974: 33 |
| BILLARD A. 1913: 17 |
| BALE W. M. 1884: 124 |
| BUSK G. 1852: 401 |