
Gekko lauhachindai, Panitvong, Nonn, Sumontha, Montri, Konlek, Kanokorn & Kunya, Kirati, 2010
|
publication ID |
https://doi.org/10.5281/zenodo.199141 |
|
DOI |
https://doi.org/10.5281/zenodo.5688408 |
|
persistent identifier |
https://treatment.plazi.org/id/038E2E37-FFA4-FFBD-FF11-FA18FA0FFB3F |
|
treatment provided by |
Plazi |
|
scientific name |
Gekko lauhachindai |
| status |
sp. nov. |
Gekko lauhachindai sp. nov.
Figures 1–3 View FIGURE 1 View FIGURE 2 View FIGURE 3
Holotype: THNHM 15903 [formerly Montri Sumontha field series (MS) 151], adult male; Thailand, Saraburi Province, Chalermphrakiat District, Sub Cha-om Cave (14°43.213’N, 100°51.209’E); collected by Nattakan Suttanon and Wut Taksintum, 23 June 2009 GoogleMaps .
Paratypes: CUMZ R 2009,6,24-2 [formerly MS 152], adult female ; PSUZC-RT 2010.50 [formerly MS 157], adult male ; ZMKU Rep-000311 [formerly MS 153], adult male, same locality and collector data as holotype GoogleMaps ; KZM 0 0 3 [formerly MS 156], adult male, locality and collector unknown .
Diagnosis. A medium-sized Gekko , snout-vent length at least 98 mm. Dorsum with 14 rows of prominent, small, smooth tubercles. Rostral approximately two times wider than deep, with I-shaped rostral groove, in contact with nostril rim. Precloacal pores in a continuous series of 12–14, femoral pores absent. Digit I and IV of pes with 13 and 13–15 enlarged subdigital scansors, respectively. Dorsal pattern of large bright spots that may be expanded to 5–6 narrow whitish crossbands intersected by a bright paravertebral dotted line from nape to sacrum. Iris color in life greenish gray or coppery brown with coppery brown around pupil margins.
Description of holotype. Adult male, SVL 79.2 mm ( Figs. 1–3 View FIGURE 1 View FIGURE 2 View FIGURE 3 ). Head long (HeadL/SVL ratio 0.30), relatively broad (HeadW/HeadL ratio 0.68), somewhat depressed (HeadH/HL ratio 0.45), distinct from neck. Lores and interorbital region slightly inflated. Snout short (SnEye/HeadL ratio 0.37), 1.25 times eye diameter (SnEye /OrbD ratio 1.28); scales on snout and forehead small in size, granular and slightly domed; scales on snout much larger than those on interorbital region. Eye relatively large (OrbD/HeadL ratio 0.29); pupil vertical with crenelated margins; supraciliaries short, posteriormost bearing minute spines. Ear opening oval, vertically oriented, small (EarL/HeadL ratio 0.06); eye to ear distance only slightly less than diameter of eyes (EyeEar/OrbD ratio 0.98). Rostral approximately 2 times wider (3.51 mm) than high (1.80 mm), “ I ” shaped median groove present; two much enlarged supranasals incompletely separated by a single anterior and two posterior, small, oval internasals; rostral in contact with supralabial I, supranasals and nostril; internasal separated from rostral by narrow anterior contact of supranasals; nostrils round, each surrounded by rostral, supranasal, first supralabial and two enlarged postnasals which enter the nostril forming a recessed valvular flange in its posterior third; 3 rows of scales separate orbit from supralabials. Mental triangular, somewhat deeper (1.26 mm) than wide (3.01 mm) and not much deeper than infralabials; one pair of elongated postmentals (2.78 mm long, 1.20 mm wide) meeting at a point behind the mental, each postmental bordered anterolaterally by first infralabial, anteromedially by a mental, medially by the other post mental, laterally by the first scale in a row of enlarged scales bordering the infralabials, and posteriorly by four small (2 times the size of a granular gular scale) chin shields, the medial one in shared contact with both left and right postmentals. Infralabials bordered by a row of enlarged scales, decreasing in size posteriorly. Enlarged supralabials to midpoint of orbit 9 (left) and 10 (right); supralabials to angle of jaws 12 (left)–13 (right); enlarged infralabials 11 (left and right); interorbital scale rows across narrowest point of frontal 24, between supraciliaries 40. Supraorbital scales highly heterogeneous in size, largest at medial edge of orbit in midorbital position.
Body robust, trunk relatively long (TrunkL/SVL ratio 0.44), dorsoventrally depressed in cross-section, with small but distinct ventrolateral folds without denticulate margins. Dorsal scales heterogeneous, granular, rectangular to oval, flattened; irregularly arranged, small (4–5 times size of granules), domed tubercles extending from posterior margin of orbit to tail; tubercles much smaller on parietal region than elsewhere, larger on mid-flanks than middorsally or on ventral flanks; tubercles in 14 rows at midbody. Ventral scales larger than dorsal, smooth, relatively hexagonal, and imbricate, largest posteriorly; somewhat larger on abdomen than on chest, becoming granular and much smaller in gular region; midbody scale rows across belly to ventrolateral folds 32. Slightly enlarged precloacal scales in a single row of 13, pitted with pores, arranged in angular series; somewhat enlarged scale rows extending posteriorly from pore–bearing scales to the anterior of cloacal lip, no enlarged femoral scales. Scales on fore and hind limbs homogeneous, smooth, flat, rounded, subimbricate without enlarged tubercles; smooth, flattened, subimbricate scales on venter of fore- and hindlimbs.
Forelimbs and hindlimbs moderately long, stout; forearm and tibia moderately long (ForeaL/SVL ratio 0.13; CrusL/SVL ratio 0.19); digits relatively short, digit I, both manus and pes, clawless, all remaining digits strongly clawed; digits of manus and pes feebly webbed at base; distal portions of digits strongly curved, arising from distal portion of expanded subdigital pad; scansors beneath each toe undivided; scansors from proximalmost at least twice diameter of palmar scales to distalmost: 11-11-16-14 -12 (right manus), 13-13-14- 15-14 (right pes). Relative length of digits of manus: IV> III> II> V> I (right); of pes: IV> V> III> II> I (right).
Tail depressed, 83% regenerated tail slightly shorter than head and body (TailL/SVL ratio 0.94); dorsal surface of original tail portion covered with small, round to oval, juxtaposed to weakly subimbricate granules forming regular transverse rows; 2 rows of round flatten tubercles, 5 times size of granules, absence of tubercles on regenerated portion, indistinct caudal segment, scales on regenerated portion much larger than on original portion; ventral caudal scales much larger, smooth, imbricate, with median enlarged subcaudal plates extending about 4/5 across the width of tail (regenerated portion). Posterior portion of dorsum of each tail segment with a single transverse row of six enlarged, rounded tubercles, each with upturned keeled or mucronate distal margin (ventrolateral pair of tubercles largest, not keeled or mucronate). One large, blunt, rounded postcloacal spur on each side of tail base.
Coloration (in preservative). Body mottled purplish brown with a series of six white T-shaped markings (due to narrow cross bars intersecting with each vertebral marking, each somewhat longer than broad) along the vertebral line from shoulder to sacrum (one on shoulder, four between axilla and groin, one across sacrum), a transverse series of white spots on nape, and a series of small white spots or narrow vertical bars near ventrolateral margin of flanks. Head mottled like dorsum with a series of whitish markings across posterior portion of parietal, three postocular stripes, one radiating nearly vertically from eye through 10th supralabial and 9th infralabial, one obliquely downward from eye to midpoint between eye and ear, and one stripe forming a roughly postparietal cross marking. Supralabials and infralabials mottled like head with narrow whitish vertical bars on edge of scales. Venter whitish, each scale with diffuse, minute dark speckles, pigmentation heaviest under parts of throat and anterior margins of hindlimbs. Limbs mottled and marked with regularly arranged whitish bars or spots. Metapodial and phalangeal joints with whitish markings, alternating with purplish-brown mottled interspaces. Regenerated tail venter more-or-less uniform pale purplish brown with darker irregular longitudinal lines.
In life body dorsum mottled golden mustard yellow on dark brown. More-or-less contiguous areas of solid purplish brown on nape and in paravertebral position. Roughly longitudinal white markings on canthal ridges. Venter grayish-white. Iris greenish gray, coppery brown around margin of pupil.
Variation. The paratypes are very similar in most respects ( Table 1 View TABLE 1 ). A female paratype (CUMZ R 2009,6,24-2) has smaller ears than all male type series (EarL/OrbD 0.16 vs 0.23–0.28), dorsum with six transverse rows of bright white spots. All male paratypes have 12 precloacal pores, absent in female. Male paratype PSUZC-RT 2010.50 has a nearly complete tail, slightly depressed without shallow groove on its dorsum; a single row of median subcaudal plates, some divided to pairs; white bars alternating with subequal purplish-brown bars, indistinct anterior borders and darker posteriorly; six white bars on original tail; venter light purplish brown, darker under chest and groin. Male paratype ZMKU Rep-000311 has six white vertebral markings and five transverse bars from nape to sacrum. Other specimens were observed during Konlek’s fieldwork: the largest individual examined, a female, had a SVL of 98 mm; some males had 14 precloacal pores; juvenile specimens exhibited the same color and pattern but with more contrast than adult specimens, with the rostral to interorbital region with olive brown overtones. A juvenile specimen had seven white bars on its original tail ( Konlek 2009) ( Fig. 5 View FIGURE 5 ).
Comparisons. Gekko lauhachindai sp. nov. may be distinguished from the large-bodied group of geckos, G. g e c k o, G. nutaphandi , G. smithii , G. s i a m e n s i s, and G. albofasciolatus Günther by its rostral participation in the nostril border, and slender body; from G. verreauxi Tytler by its smaller size (adult SVL<130); from G. subpalmatus Günther, G. m e l l i Vo g t, G. athymus Brown & Alcala , G. scientadventura Rösler et al. , and G. tawaensis Okada by the presence of dorsal tubercles; from G. v i t t a t u s Houttuyn, G. crombota Brown et al. , G. palmatus Boulenger , G. porosus Taylor , G. gigante Brown & Alcala , G. kikuchii Oshima , G. mindorensis Taylor , G. monarchus (Schlegel) , G. romblon Brown & Alcala , G. palawanensis Taylor , G. ernstkelleri Rösler et al. , G. ro s s i Brown et al., and G. carusadensis Linkem et al. by its lack of femoral pores; from G. t a w a e n s i s Okada, G. v e r t e b r a l i s Toda et al., and G. s h i b a t a i Toda et al. by the presence of precloacal pores in males; from G. japonicus (Schlegel) , G. swinhonis Günther , G. taibaiensis Song, G. auriverrucosus Zhou & Liu , G. scabridus Liu & Zhou , G. liboensis Zhou & Liu , and G. wenxianensis Zhou & Wang by its greater number of digital lamellae under toe IV (13–15 vs <9); from G. yakuensis Matsui & Okana by greater number of digital lamellae under toe I (13 vs. 10); from G. chinensis Gray and G. hokouensis Pope by its greater number of dorsal tubercle rows (14 vs 10); from G. canhi Rösler et al. by its greater number of precloacal pores (12–14 vs 5); from G. similignum Smith by fewer mid body scale rows (112–121 vs. 144–153).
Within the morpho-group of G. petricolus including G. russelltraini , G. t a k o u e n s i s, G. badenii , G. grossmanni , G. scientiadventura and G. vietnamensis , the new species can be distinguished from G. badenii by the presence of a series of vertebral whitish spots on dorsum versus rather plain yellowish and grey coloration; from G. petricolus and G. takouensis by the presence of white transverse bars on dorsum, and lower count of subdigital scansors under toe IV (13–15 vs. 20 and 18–20, respectively); from G. grossmanni by the presence of an “I-shaped” rostral groove versus a “Y” or “X” shapedgroove, presence vs. absence of white transverse bars on dorsum and vertical bars on mid-flank, and lower count of subdigital scansors under toe IV (13–15 vs. 18–20); from G. vietnamensis Nguyen by the presence of precloacal pores and from the morphologically most similar species, G. russelltraini Ngo et al. by transverse bars crossing vertebral markings (versus mid-flank vertical bars only), greater number of mid-body ventral scale rows (32 vs. 28–30), lower number of subdigital scansors on toe IV (13–15 vs. 17–19), and shorter snout (SnEye/HeadL ratio 0.35–0.39 vs 0.40–0.47), lower SnEye/OrbD ratio (1.20–1.30 vs. 1.50–1.91).
Etymology. The specific epithet is a patronym in genitive case honoring the Thai herpetologist, Associate Professor Dr. Virayuth Lauhachinda who has dedicated his life to teach many generations of herpetologists in Thailand. We suggest the following common names: Lauhachinda’s Cave Gecko (English) , Tuk Kae Thum Arjarn Virayuth (Thai).
Distribution and natural history. G. lauhachindai sp. nov. has been found in two limestone karst formations: Khao Wong and Khao E-Mod, Chalermphrakiat District, Saraburi Province, central Thailand ( Figs. 5–6 View FIGURE 5 View FIGURE 6 ). Saraburi province’s lime-stone karst formations are of Permian origin, as evidenced from fossil records of animals such as fusulinids and crinoids from that period. The karst area was formed when the subcontinental Shan-Thai collided with sub-continental Indochina, generating a karstic limestone formation in the middle of Thailand ( Witayarut 1996).
G. lauhachindai sp. nov. were found exclusively in caves and areas immediately adjacent to cave entrances, clinging to both rough and smooth surfaces and tree roots near holes and crevices, to which they would retreat if disturbed. They share this habitat with two other congeners, G. gecko and G. siamensis . The former is a widespread species that can be found in various kinds of habitats, the latter was described from Nakhon Ratchasima Province, northeast of Saraburi Province. It is not unexpected to find G. siamensis in Saraburi Province as the limestone ranges of these two adjacent provinces are somewhat connected. At the type locality where three species occur G. lauhachindai sp. nov. are confined to caves and their entrances, whereas G. siamensis can be found both in caves and in karst areas further away from cave entrances, and in absence of G. g e c k o it can also be found on large tree trunks and manmade structures near karst areas. Gekko gecko can normally be found further away from caves on isolated limestone outcrops, large tree trunks and manmade structures.
Other gekkonids found in and around the area are Cyrtodactylus chanhomeae Bauer et al. , Hemidactylus frenatus (Schlegel) , H. platyurus (Schneider) , Gehyra fehlmanni (Taylor) , Hemiphyllodactylus typus Bleeker , Dixonius melanostictus (Taylor) , and D. siamensis (Boulenger) . Both Hemidactylus spp. are more adapted to human disturbance and found mostly on manmade structures and tree trunks both in and away from karst areas. Gehyra fehlmanni was found exclusively on smooth limestone walls in and around cave entrances. Cyrtodactylus chanhomeae are confined to caves and their entrances, their behavior and habitat being very much the same as G. lauhachindai sp. nov. Dixonius melanostictus preferred areas outside caves, foraging under leaf litter, and on limestone outcrops not far from the ground level. A single D. siamensis was observed once in the area but this species normally lives further away from karst areas. A one year survey in the area by the third author also recorded a single Hemiphyllodactylus typus which was found on limestone outcrop not far away from cave entrance.
Females carrying eggs were observed during January and May, indicating that the gecko lays eggs during the dry season, however oviposited eggs have not yet been found. Young animals, SVL 34 mm and 38 mm, were observed in April and May, respectively. Feces of the newly captured paratype CUMZ R 2009,6,24-2- were examined by Paradorn Dokchan of the Environmental Entomology Research and Development Center, Kasetsart University (Kamphaeng Saen Campus), and revealed four orders of insects, Hemiptera represented by four families – Delphacidae , Cicadellidae and two unidentifiable, Coleoptera represented by one unidentifiable family and Coccinellidae [ Menochilus sexmaculatus (Fabricius) ], Odonata represented by the Coenagrionidae , and Blattodea represented by the family Blattellidae . Shed skin was also recovered from the alimentary tract.
Potential predators in the area are domestic cats and snakes. The former are quite common due to the proximity of human settlements (e.g. temples and houses). We recorded four species of snakes in the cave area, Cryptelytrops albolabris (Gray) , C. macrops (Kramer) , Bungarus candidus (Linnaeus) , and Dryocalamus davidsonii (Blanford) . Interestingly, on one occasion, we observed adult Gekko siamensis eating adult Dixonius melanostictus , representing the common phenomenon of larger gekkonids preying upon smaller species.
The closest relative of G. lauhachindai sp. nov. that also occurs in Thailand is G. petricolus , which is known from no closer than the Ta Phraya District of Sa Kaeo Province, ( Nabhitabhata et al. “2000 ” 2004), approximately 200 kilometers southeast of Saraburi Province.
Dorsal scale scale Number scansors IV SVL Mid Mid rows toe
G. lauhachindai sp. nov. 98.0 14 32 112–121 12–14 13–15 11–12 11 G. badenii Shcherbak&Nekrasova, 1994 108.0 8–13 29 –35 114–136 10–18 15–20 11–15 9–14 G. grossmanni Günther, 1994 89.0 presence 26–31 116–135 8–14 18–20 11–13 9–11 G. petricolus Taylor, 1962 98.0 16 27–32 <161 9–10 20 12 10–11 G. russelltraini Ngo et al., 2009 82.9 12–16 28–30 90–107 8–11 17–19 12–15 11–14 G. scientiadventura Rösler et al., 2005 73.0 absence 38–43 118–140 5–8 14–17 12–14 9–13 G. ta k o ue n si s Ngo&Gamble, 2010 107.0 14–17 30–34 83–93 11–14 18–20 13–16 10–12 G. vietnamensis Ngunyen, 2010 91.0 presence n/a 28–30 0 18–20 11–12 10 –11


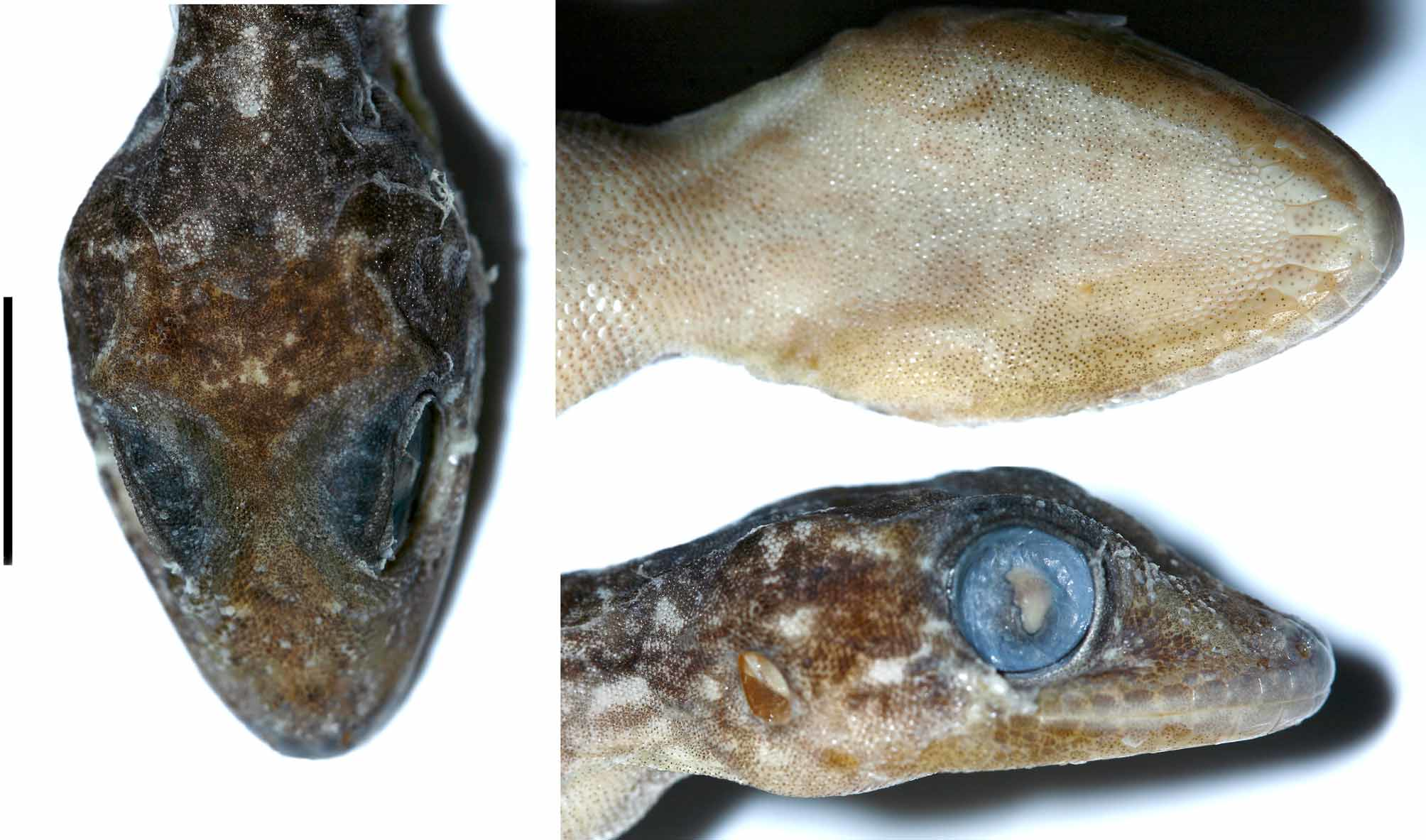

TABLE 1. Mensural and meristic data for the type series of Gekko lauhachindai sp. nov. Abbreviations as in Materials and Methods, all measurements in mm.
| THNHM 15903 Holotype | CUMZ R 2009,6,24-2 Paratype | PSUZC-RT 2010.50 Paratype | ZMKU Rep- 0 0 0 311 Paratype | KZM 0 0 3 Paratype | |
|---|---|---|---|---|---|
| Sex | Male | Female | Male | Male | Male |
| SVL | 79.2 | 78.9 | 81.2 | 78.5 | 78.0 |
| ForeaL | 10.4 | 10.2 | 9.8 | 10.8 | 9.7 |
| CrusL | 15.4 | 14.4 | 16.1 | 15.1 | 14.4 |
| TailL | 74.4 (regen. 62.1) | 71.2 (regen. 28.3) | 76.5 (regen. 13.1) | 74.8 (regen. 63.3) | 63.7 (regen. 54.7) |
| TailW | 9.0 | 7.8 | 7.2 | 7.8 | 8.3 |
| TrunkL | 34.7 | 37.5 | 36.1 | 34.7 | 37.0 |
| HeadL | 23.7 | 22.4 | 23.7 | 23.7 | 23.3 |
| HeadW | 16.2 | 15.5 | 15.6 | 15.9 | 16.1 |
| HeadH | 10.7 | 9.8 | 9.6 | 9.9 | 9.2 |
| OrbD | 6.8 | 6.9 | 6.9 | 6.8 | 7.1 |
| EyeEar | 6.6 | 6.1 | 5.6 | 6.2 | 6.1 |
| SnEye | 8.7 | 8.7 | 8.3 | 8.9 | 8.5 |
| NarEye | 7.1 | 6.3 | 6.8 | 7.0 | 6.7 |
| Interorb | 7.4 | 8.2 | 7.9 | 8.9 | 9.4 |
| EarL | 1.5 | 1.1 | 1.7 | 1.9 | 1.9 |
| Internar | 3.0 | 2.4 | 3.4 | 2.9 | 2.6 |
| Tubercle Rows | 14 | 14 | 14 | 14 | 14 |
| Ventral Scales | 32 | 32 | 32 | 32 | 32 |
| Precloacal pores | 13 (pits) | - | 12 (pits) | 12 (pits) | 12 (pits) |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Gekko lauhachindai
| Panitvong, Nonn, Sumontha, Montri, Konlek, Kanokorn & Kunya, Kirati 2010 |
G. vietnamensis
| Ngunyen 2010 |
G. russelltraini
| Ngo et al. 2009 |
G. scientiadventura Rösler et al., 2005
| Rosler et al. 2005 |
G. badenii
| Shcherbak&Nekrasova 1994 |
G. grossmanni Günther, 1994
| Gunther 1994 |
G. petricolus
| Taylor 1962 |