
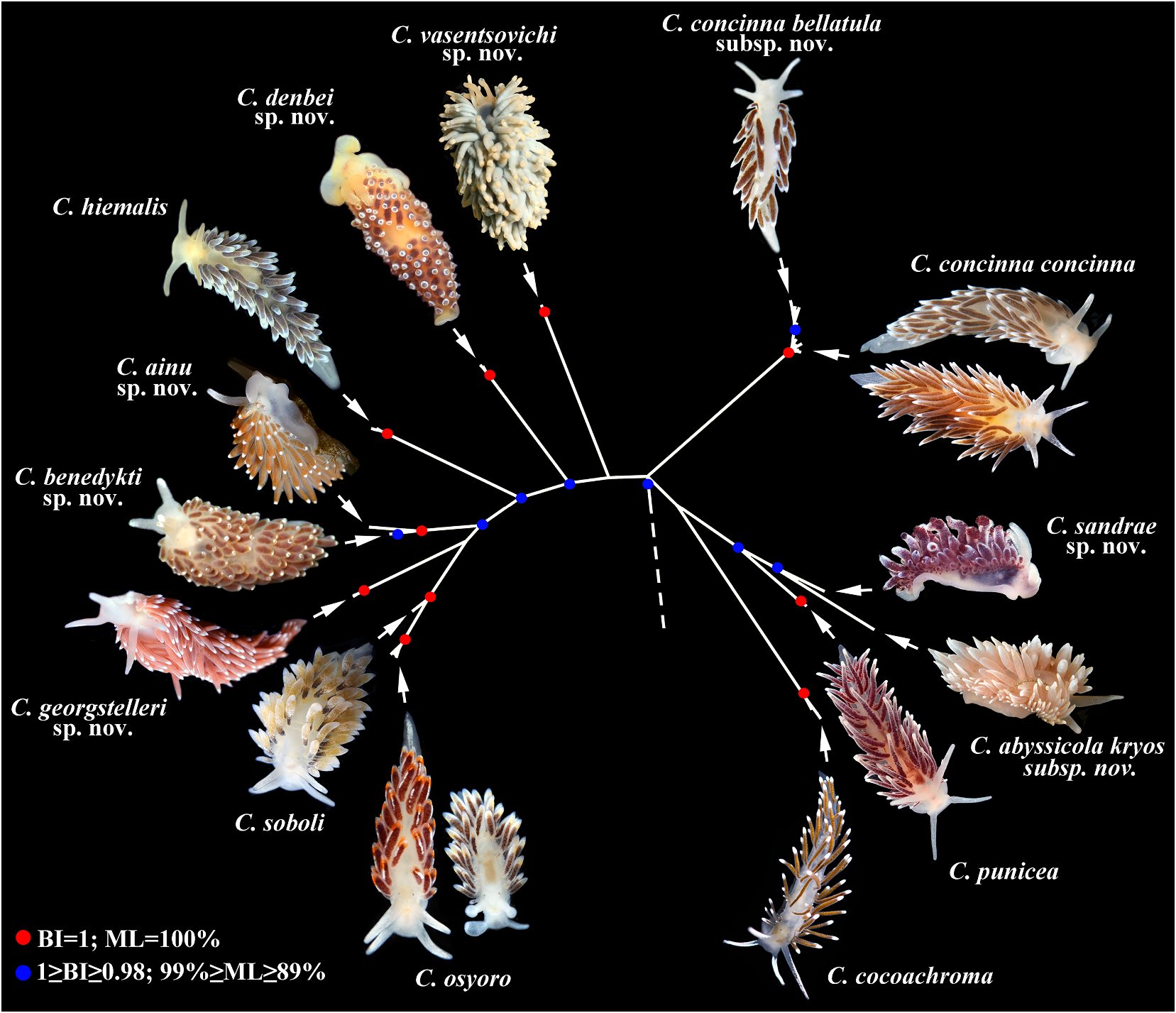
Cuthonella ainu, Korshunova & Sanamyan & Sanamyan & Bakken & Lundin & Fletcher & Martynov, 2021
|
publication ID |
https://doi.org/ 10.1163/18759866-BJA10017 |
|
persistent identifier |
https://treatment.plazi.org/id/038C8782-E475-9265-FF7E-FA05FDBE4EFA |
|
treatment provided by |
Felipe |
|
scientific name |
Cuthonella ainu |
| status |
sp. nov. |
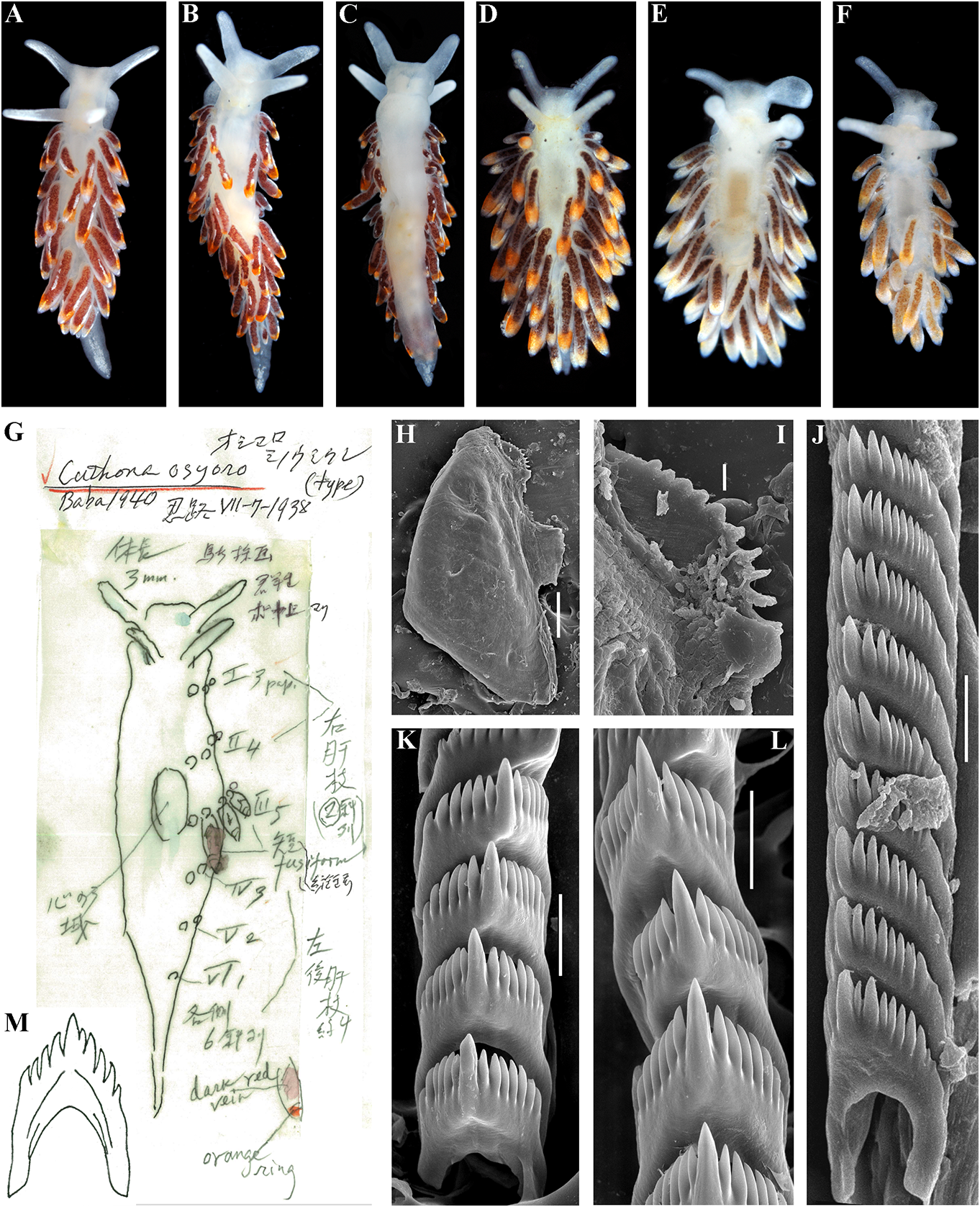
Cuthonella ainu View in CoL sp. nov. Etymology. Named in honour of the Ainu,
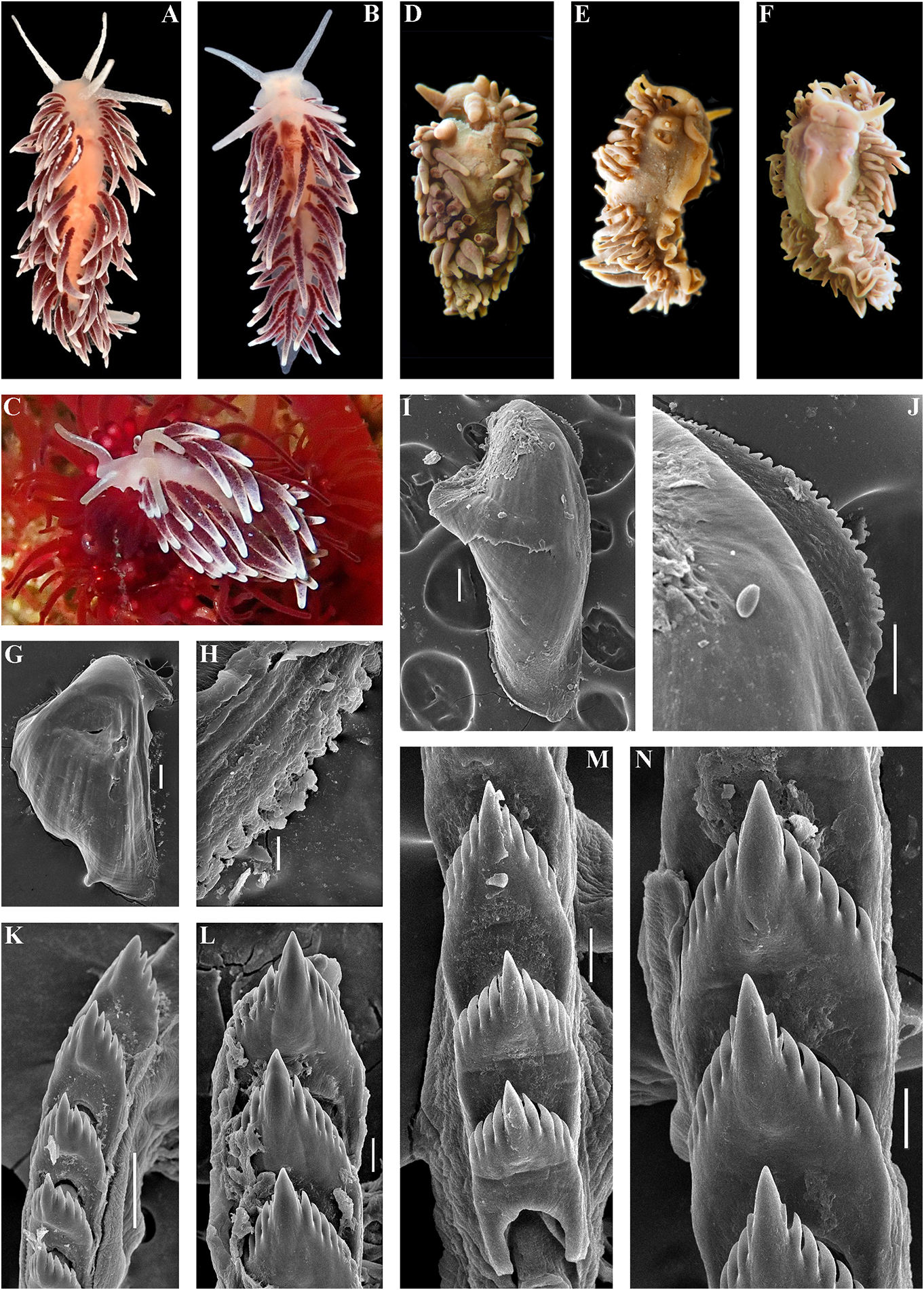
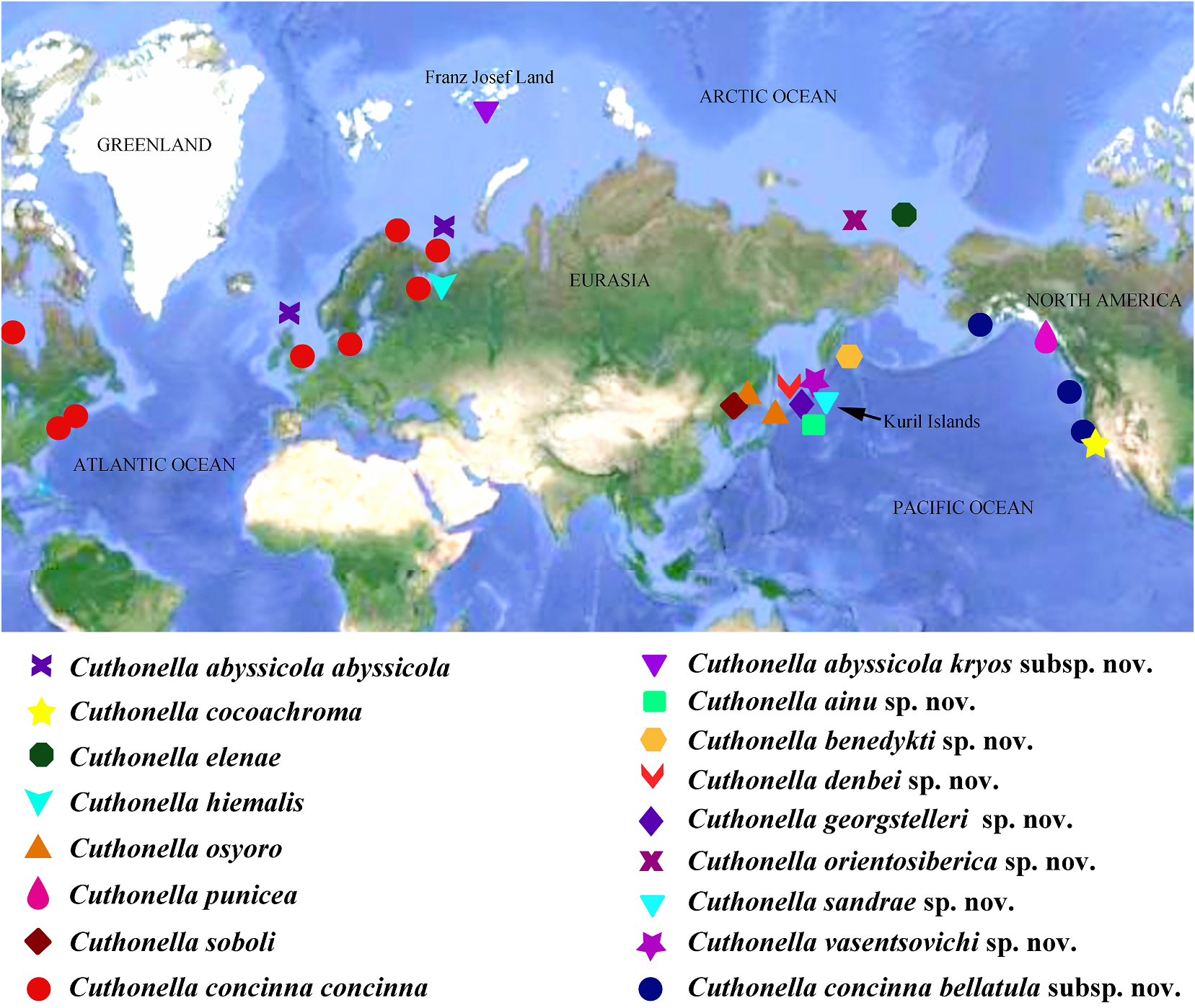
( Figs 1 View FIGURE 1 , 2 View FIGURE 2 , 6 View FIGURE 6 , 19C View FIGURE 19 , 21 View FIGURE 21 ) the indigenous people of the Kuril Islands
ZooBank: http://urn:lsid:zoobank. (type locality of this new species), southern o r g:a c t:2 9D B6C 9 4 - 4 5E 8 - 4 8 2 6- 9 0 7 D - Kamchatka, Sakhalin, Hokkaido and northern
2F5B53B85413 Honshu (Tohoku). The Ainu speak a highly
Type material. Holotype. ZMMU Op-618, isolated language, which is now critically
1 spec., L = 4 mm length (preserved), Pacific endangered. The Ainu were forced to leave
Ocean, Middle Kuril Islands, Matua Island, their original lands both by Japanese and
Klyuv Cape, 14 m, collected by N.P. Sanamyan, Russian Downloaded authorities from. Ainu Brill. com de facto 12/12was /2023extinct 04:16:22PM
04.08.2017. via Open Access. This in Russia is an open since access the article early distributed 70s, but attempts under the to terms of the CC-BY 4.0 license. https://creativecommons.org/licenses/by/4.0/
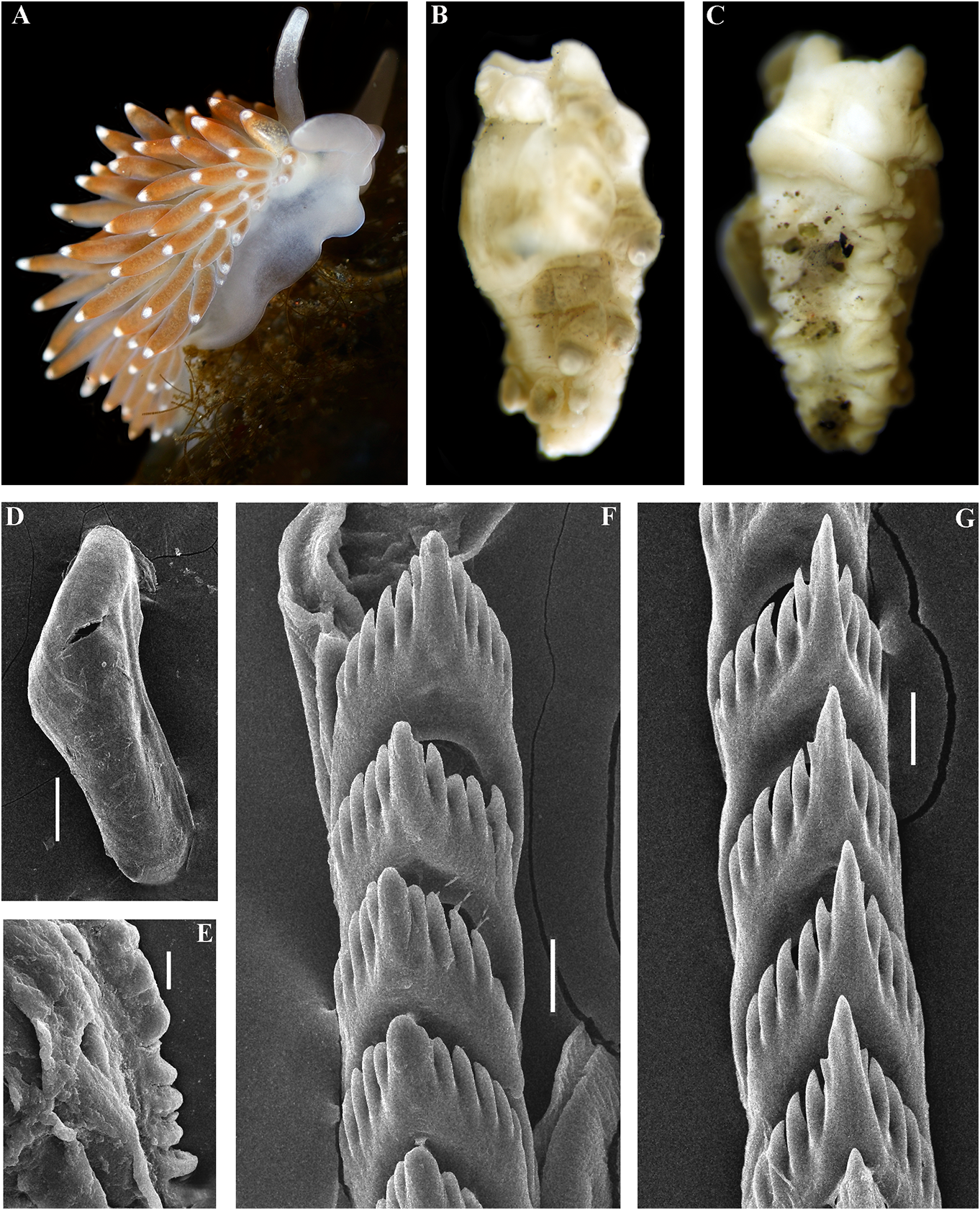
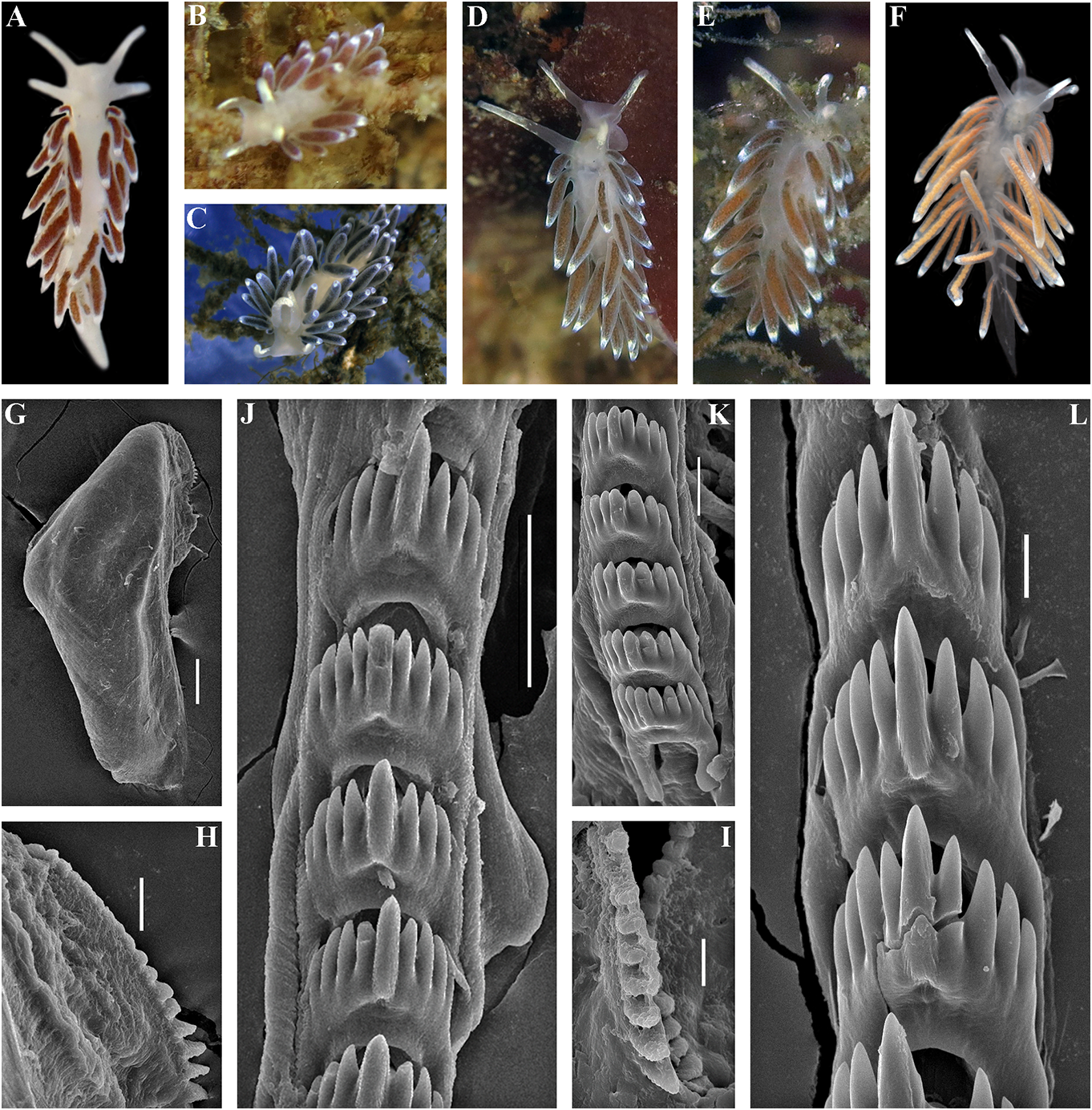
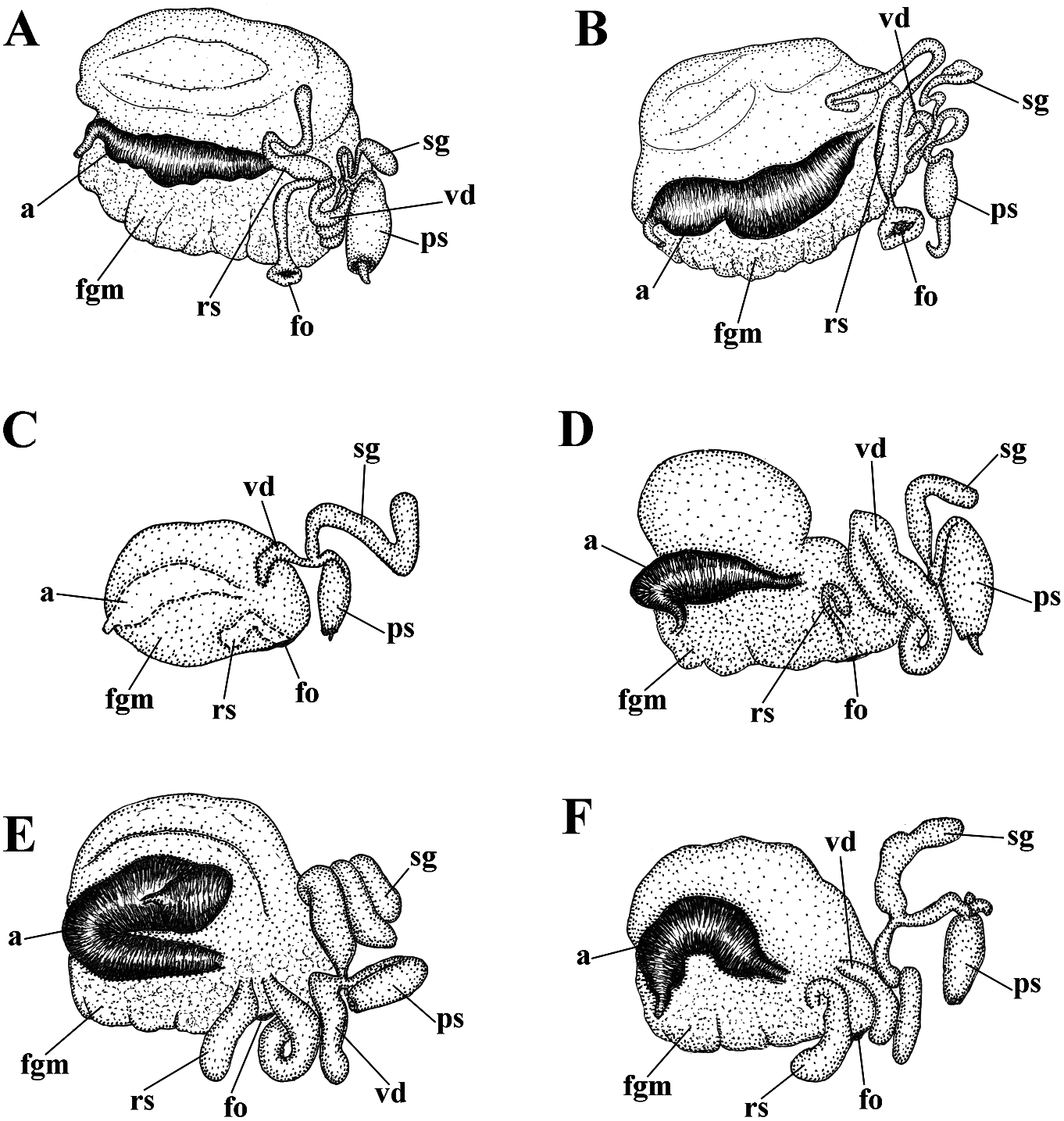
revitalize Ainu culture based on descendants white spots. Rhinophores semi-transparent withpartialAinuancestryhavebeencarriedout and covered with a dense white pigment at recently in Kamchatka without governmental the top (fig. 6A). support and official recognition (e.g., Tanaka, Jaws. Jaws broad, yellowish in colour. 2017). In Japan, formerly forcedly-assimilated Masticatory processes of jaws covered with a Ainu now have governmental support for their single row of distinct denticles (fig. 6D, E). language and culture with official recognition Radula. Radula formula 18 × 0.1.0. Central ( Tsunemoto, 2019). tooth elongated with a strongly protracted, External morphology. Body moderately pointed non-compressed cusp (fig. 6F, G). wide. Foot and tail broad. Oral tentacles Central tooth bears up to 7 lateral denticles. moderate. Rhinophores similar in size to oral Cusp clearly delineated from the adjacent first tentacles, smooth to slightly wrinkled. Dorsal lateral denticles. cerata elongate, thick, arranged in a moderate Reproductive system. Diaulic. Hermanumber of continuous rows. Five pre-anal phroditic duct leads to a relatively short, unbranched ceratal rows. Anal opening slightly bent, swollen ampulla. Vas acleioproctic. deferens short, without a distinct prostate. Colour (live). Background colour yellowish Supplementary gland long, inserts into the vas white. Digestive gland diverticula in cerata deferens a short distance from the elongated brownish to light orange. Ceratal tips with penial sheath Downloaded. Penis conical from Brill.com, unarmed 12/12/2023. 04:16:22PM opaque white spot, sometimes with very via Open small Oviduct Access. This is an connects open access through the article distributed insemination under the terms of the CC-BY 4.0 license.
https://creativecommons.org/licenses/by/4.0/
duct into the female gland complex. where he started his research (Wszolek et al., Receptaculum seminis in distal position, on a 1990). He also served as the only physician short stalk, elongate-oval (fig. 19C). at the time (starting in 1879) for the large Ecology. Only found on soft bottom with region of Kamchatka, the type locality of this stones at 14 m depth. new species. Among other achievements, Distribution. Matua Island, Middle Kuril Dybowski contributed to Kamchatka’s Ainu Islands. dictionary.
Remarks. According to the molecular External morphology. Body moderately phylogenetic analysis Cuthonella ainu sp. nov. is wide. Foot and tail broad. Oral tentacles closely related to C. benedykti sp. nov. (figs 1, 2). moderate. Rhinophores blunt and similar The maximum intragroup distance in C.ainu sp. in size to oral tentacles, smooth to slightly nov.is0.15%.The lowest COI intergroup distance wrinkled. Dorsal cerata elongate, thick,
of 2.65% is found between C. ainu sp. nov. and arranged in continuous rows. Five pre-anal C. benedykti sp. nov. (table 1). Morphological unbranched ceratal rows. Anal opening analysis reveals differences in radular patterns acleioproctic.
between C.benedykti sp. nov. and C.ainu sp. nov. Colour (live). Background colour yellowish (figs 6F, G and 7G, H). Externally, C. ainu sp. nov. white. Digestive gland diverticula in cerata differs from C. benedykti sp. nov. by its orange- brownish with a darker reddish to brown brownish colouration and spot-shaped rather hue. Ceratal tips with opaque white ring or than ring-shaped patterns of the white pigment spot, sometimes with additional yellow ring, on the ceratal apices. For a morphological and small white spots. Rhinophores light comparison with other Cuthonella species, see yellowish with white pigment on the top (fig.
table 2. 7A–D).
Jaws. Jaws broad, yellowish in colour.
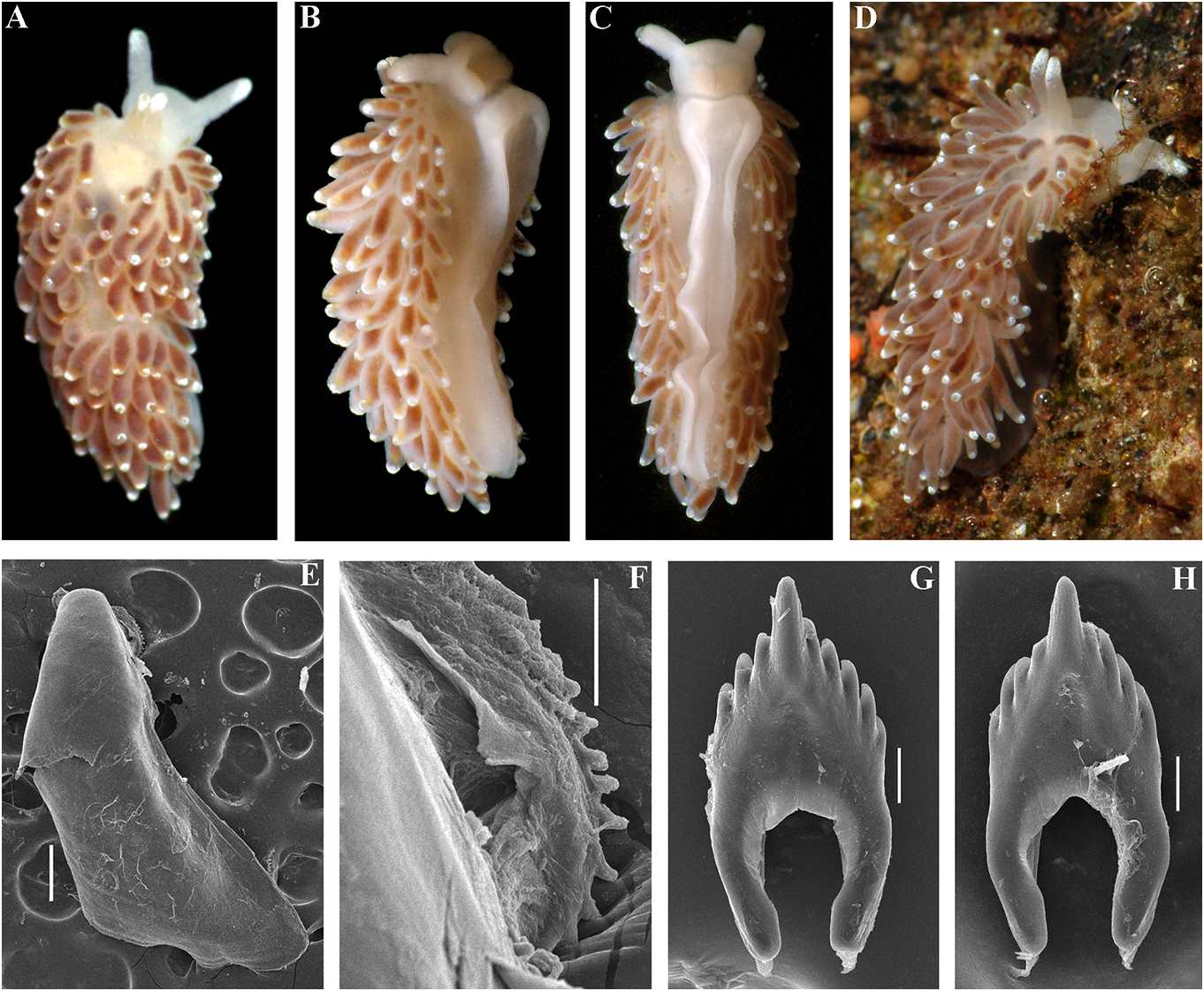
Cuthonella benedykti sp. nov. Masticatory processes of jaws covered with a
( Figs 1 View FIGURE 1 , 2 View FIGURE 2 , 7 View FIGURE 7 , 19D View FIGURE 19 , 21 View FIGURE 21 ) single row of distinct denticles (fig. 7E, F).
ZooBank: http://urn:lsid:zoobank. Radula. Radula formula 36 × 0.1.0
o r g:a c t:D4 C D E 4F 2-0 5F 3- 4 6E 4 - 8C6 B - (specimen 6 mm in length). Central tooth B233722AAB25 elongated with a strongly protracted, pointed Type material. Holotype. ZMMU Op-194, non-compressed cusp (fig. 7G, H). Central
1 spec., L = 6 mm length (preserved), Pacific tooth bears up to 5 lateral denticles. Cusp Ocean, Kamchatka, Starichkov Island , 6–12 m, clearly delineated from the adjacent first stones, collected by T. A. Korshunova and A. V. lateral denticles .
Martynov , 19.08.2008. Reproductive system. Diaulic. Herma- Etymology. The name is in honour of a phroditic duct leads to a relatively short, polymath scientist of Polish origin, Benedykt swollen ampulla. Vas deferens wide and Dybowski (1833–1930), who made substantial moderate in length, without a distinct achievements especially in zoology and the prostate. Supplementary gland relatively anthropology of Russian Siberia and the Far short, inserts into the vas deferens a short East. Benedykt Dybowski belonged to the distance from the elongated penial sheath.
Polish intelligentsia, and he was arrested after Penis conical, unarmed. Oviduct connects the Polish rebellion against the Russian Empire through the insemination duct into the female (1863, “January Uprising”) and sentenced gland complex. Receptaculum seminis in to death, later changed to exile in Siberia, distal position Downloaded, on from a short Brill.com stalk12, oval /12/2023 (fig. 04 19:16 D:22). PM via Open Access. This is an open access article distributed under the terms of the CC-BY 4.0 license. https://creativecommons.org/licenses/by/4.0/ Ecology. Lives on stones at depths of c. feature needs to be particularly commented 6– 12 m. on, because all the other 15 species of the genus Distribution. Kamchatka, Pacific side. Cuthonella invariably possess a supplementary Remarks. According to the molecular gland inserted into the vas deferens and not phylogenetic analysis Cuthonella benedykti into the penial sheath or penial base. The sp. nov. forms a separate clade sister to C. ainu reproductive system of C. cocoachroma (not sp. nov. (figs 1, 2). The maximum intragroup available for the present study) needs to be distance in C. benedykti sp. nov. is 0.16%. The re-examined in this respect, considering that minimum COI intergroup distance of 2.65% according to the molecular data it belongs to the is found between C. benedykti sp. nov. and C. genus Cuthonella (fig. 1). The thecate hydroid ainu sp. nov. ( Table 1). Morphological analysis Thuiaria sp. was reported as a food object for C. reveals considerable differences in the cocoachroma (Goddard, 1985) . The maximum denticulation of the central teeth between C. intragroup distance in C. cocoachroma is benedykti sp. nov. and its sister C. ainu sp. nov. 1.52%. Interestingly, the minimum COI as well as more distantly related, C.georgstelleri intergroup distance of 9.74% is found between sp. nov., C. hiemalis , C. osyoro , and C. soboli C. cocoachroma and the allopatric C. punicea (figs 6, 7, 11, 12, 14, 17). Externally, C. benedykti (table 1) rather than between the sympatric C. sp. nov. (fig. 7A–D) differs from C. ainu sp. nov. cocoachroma and C. concinna bellatula subsp. (fig. 6A) by a more brownish colouration with nov. which are externally nearly identical. a dark reddish hue, and more distinct ring- For a morphological comparison with other shaped white pigment on the ceratal apices. Cuthonella species, see table 2. For a morphological comparison with other Cuthonella species, see table 2. Cuthonella concinna concinna (Alder &
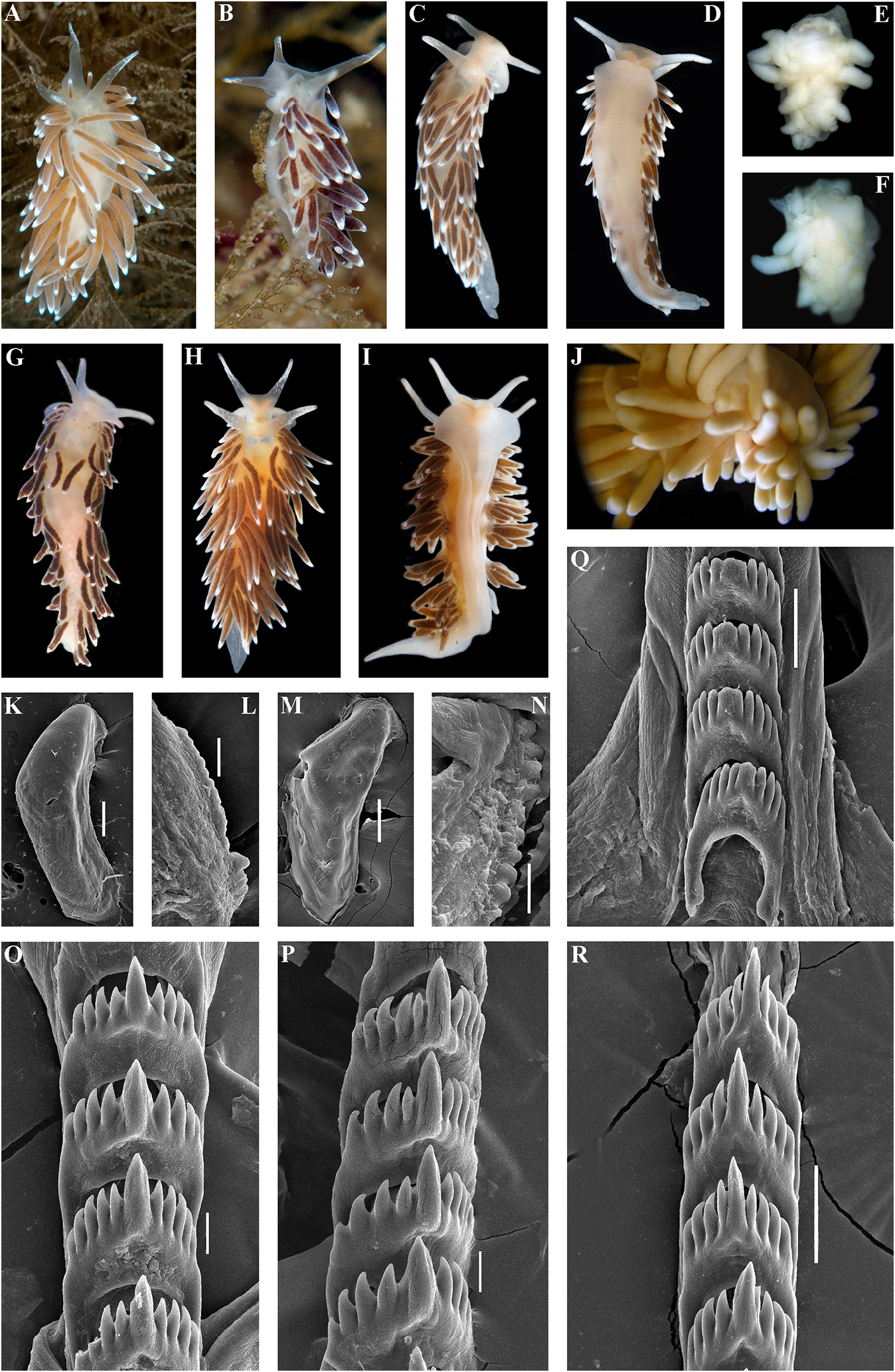
Hancock, 1843) Cuthonella cocoachroma (Williams & ( Figs 1–3 View FIGURE 1 View FIGURE 2 View FIGURE 3 , 8 View FIGURE 8 , 19E View FIGURE 19 , 21 View FIGURE 21 )
Gosliner, 1979) Eolis concinna Alder & Hancock, 1843: 234 ; Cuthona cocoachroma Williams & Gosliner, Alder & Hancock, 1845 –1855: 50, fam. 3, pl. 24,
1979: 204–211, figs. 3–6. plus unnumbered pages. Remarks. Cuthonella cocoachroma has been Cuthona concinna – Brown, 1980: 249–250, described in detail by Williams & Gosliner fig. 5H; 6D, H. (1979),which will not be repeated here except to Cuthona distans Odhner, 1922: 28–29 , figs 11, say that it differs from the sympatric C.concinna 12, syn. nov. bellatula in having rhinophores which are twice Cratenopsis concinna – Nordsieck, 1972: 81. as long as the oral tentacles, having fewer pre- Trinchesia concinna – Pruvot-Fol, 1954: 385. anal ceratal rows (3–4 vs 5–6) and fewer ceratal Cuthonella marisalbi Roginskaya, 1963 : rows altogether (9–10 vs 10–12), and lacking 258–264, figs 1–7; Martynov & Korshunova, scattered white or bluish spots on its cerata. 2011: 200 –201, figs. In the present material commonly up to 4 pre- Cuthonella concinna – Martynov , 1992: 23; anal rows in C. concinna bellatula , that make Martynov , 2006:290; Martynov & Korshunova, external differences from C. cocoachroma as 2011: 198 –200, figs. more subtle. According to figure 5D in Williams Type material. Natural History Museum, & Gosliner (1979), C. cocoachroma possess a London, one specimen collected by Joshua supplementary gland which is inserted into Alder at Cullercoats (original type locality) the penial base instead of the vas deferens. This and indicated Downloaded as “ from type ” (NHM Brill.com, 2020). 12/12/2023 04:16:22PM
via Open Access. This is an open access article distributed under the terms of the CC-BY 4.0 license.
https://creativecommons.org/licenses/by/4.0/ Other material. ZMMU Op-41, 2 spec., Berlevåg, 70.85021°N 29.151252°E, intertidal GoogleMaps ,
L = 9 mm and 5 mm length (preserved), Barents collected by T.A. Korshunova, A.V Martynov ,
Sea, Dalnie Zelentsy,intertidal,collected byT. A. and K. Hårsaker, 01.05.2019. NTNU-VM 76044 View Materials , Korshunova, 08.2006. KM 452 , L = 3 mm length 1 spec., Finnmark, KamØyvaer, 71.049696°N (preserved), Barents Sea, Dalnie Zelentsy , 0.2– 25.91252°E, intertidal, collected by T. Bakken GoogleMaps ,
0.5 m depth, collected by T . A. Korshunova and C. Skauge, 05.05.2019. NTNU-VM 76045 View Materials , 2 View Materials
A. V. Martynov , 05.08.2005 . KM 453 , L = 8 mm spec., Finnmark, KamØyvaer, 71.049696°N length (preserved), Barents Sea, Dalnie 25.91252°E, intertidal, collected by GoogleMaps T. Bakken, Zelentsy, 0.2–0.5 m depth, collected by T.A. C. Skauge, 05.05.2019 . NTNU-VM 75960 View Materials , 1 View Materials Korshunova and A . V. Martynov , 01.08.2005 . spec., Finnmark, BugØynes , 69.972744°N ZMMU Op-523, two spec., L = 7 mm and 8 mm 29.627895°E, intertidal, collected by T.A.
length (preserved), White Sea, Rugozerskaya Korshunova and A.V. Martynov , 29.04.2019.
Bay, intertidal, collected T.A. Korshunova and NTNU-VM 75981, 1. spec., Finnmark, Berlevåg,
A. V. Martynov , 15.06.2009 . GNM Gastropoda 70.850088°N 29.154291°E, intertidal, collected 8866, L = 9.5 mm length (preserved), by GoogleMaps T. Bakken , T.A. Korshunova, A . V. Martynov , Skagerrak , Sweden, 5–10 m depth, collected and K. Hårsaker, 30.04.2019 . NTNU-VM 75992 View Materials ,
by Klas Malmberg. KM 616 , L = 3.5 mm length 1. Spec., Finnmark, Berlevåg, 70.85021°N (preserved), the North Atlantic, Northern 29.151252°E, intertidal, collected by T GoogleMaps .A.
Ireland, Strangford Lough, UK, 10–20 m Korshunova , A. V Martynov , and K. Hårsaker, depth, collected B. Picton, 23.05.2015. ZMO 01.05.2019. NTNU-VM 75994 View Materials , 4 View Materials . Spec .,
D 25946 (lectotype of “ Cuthona” distans ), Finnmark, Berlevåg , 70.85021°N 29.151252°E GoogleMaps ,
L = 3 mm length (preserved), Finnmark, intertidal, collected by T. A. Korshunova, A. V VadsØ, collected G.O. Sars. KM 767 , 1 spec., Martynov , and K. Hårsaker, 01.05.2019 .
L = 12.5 mm (live), Finnmark, Kirkenes vicinity, External morphology. Body moderately wide.
intertidal, collected by T.A. Korshunova and Foot and tail broad. Rounded foot corners
A. V. Martynov , 29.04.2019 . NTNU-VM 76078 View Materials , present.Oral tentacles moderate.Rhinophores
1 spec., Finnmark, Altafjorden , 70.156664°N similar in size, or slightly longer than oral 23.285329°E, intertidal, collected by T.A. tentacles, smooth to slightly wrinkled. Dorsal Korshunova, A. V Martynov , and K. Hårsaker, cerata elongate, arranged in continuous rows GoogleMaps .
08.05.2019. NTNU-VM76079 View Materials , 1spec., Finnmark, Up to 6 pre-anal unbranched ceratal rows .
Altafjorden, 70.156664°N 23.285329°E, Anal opening acleioproctic.
intertidal, collected by T.A. Korshunova, Colour (live). Background colour yellowish
A. V Martynov , and K. Hårsaker, 08.05.2019. white. Digestive gland diverticula in cerata NTNU-VM 75959 View Materials , 1 spec., Finnmark, BugØynes, brown to purplish-brown. Ceratal tips 69.972744°N 29.627895°E, intertidal, collected with opaque white subapical band or spot GoogleMaps ,
by T.A. Korshunova and A.V. Martynov , sometimes with a bluish hue and smaller 29.04.2019.NTNU-VM76009, 1spec., Finnmark, white spots. Rhinophores and oral tentacles Porsanger, 70.245495°N 24.999314°E,intertidal, semi-transparent with commonly white collected by T.A. Korshunova, A.V Martynov , pigment on the tips (fig. 8A–D, G–I).
and K. Hårsaker, 03.05.2019. NTNU-VM 76010, Jaws. Jaws broad, yellowish in colour.
1 spec., Finnmark, Porsanger, 70.245495°N Masticatory processes of jaws covered with a 24.999314°E, intertidal, collected by T.A. single row of distinct denticles (fig. 8K–N) GoogleMaps .
Korshunova, A.V Martynov , and K. Hårsaker, Radula. Radula formula30× 0.1.0(specimen 03.05.2019.NTNU-VM 75991, 1.spec., Finnmark, 8 mm in length Downloaded), 29 × from0.1Brill.0 (specimen.com 12/12/2023 9.5 mm 04:16:22PM via Open Access. This is an open access article distributed under the terms of the CC-BY 4.0 license. https://creativecommons.org/licenses/by/4.0/ in length), 19×0.1.0 (specimen 3 mm in length), In this study we also obtained recently 26 × 0.1.0 (specimens 8 mm in length). Central collected specimens (fig. 8C, D) from the tooth elongated with a strongly protracted type locality of the enigmatic Norwegian cusp (fig. 8O–R). Central tooth bears up to 10 species Cuthona distans Odhner, 1922 in the lateral denticles (more commonly, up to 4–5). northernmost mainland Norwegian county of Cusp clearly delineated from the adjacent first Finnmark. We also studied the type specimen lateral denticles. of C. distans from the Oslo Natural History Reproductive system. Diaulic. Museum in Norway (ZMO D 25946, designated Hermaphroditic duct leads to a relatively here as the lectotype) (fig. 8E, F). According to short or folded swollen ampulla. Vas deferens the morphological study of the type material relatively long, without a distinct prostate. of C. distans , it has a supplementary gland that Supplementary gland usually relatively long, inserts into the vas deferens relatively close sometimes shorter, inserts into the vas deferens to the penis, similar to that of C. concinna at a relatively short distance from the elongated concinna . Molecular analysis shows that penial sheath. Penis conical, unarmed. Oviduct new specimens from Finnmark belong to C. connects through the insemination duct into concinna concinna (figs 1–3). Morphological female gland complex. Receptaculum seminis data from the original description of C. in a more proximal position than in most distans ( Odhner, 1922) and our study of the other Cuthonella species, and on a short stalk, type material also confirm that C. distans without a distinct reservoir (fig. 19E). syn. nov. is a junior synonym of C. concinna Ecology. Lives on stones and rocks. concinna . In the northern localities such as Intertidal to 30 m depth. the Barents Sea and the Finnmark region, C. Distribution. Boreal European waters of the concinna concinna is one of the most common NE Atlantic ( France, British Isles, Netherlands, shallow-water species of nudibranchs. Several Denmark, Sweden, Norway to the Barents Sea thecate hydroid species of the families and the White Sea in Russia) and the Atlantic Campanulariidae and Sertulariidae were coast of the USA and Canada. reported as food objects for C. concinna Remarks. In the present study we obtained concinna : Diphasia fallax (Johnston, morphological and molecular data from 1847), Dynamena pumila (Linnaeus, 1758) , specimens of C. concinna concinna from the Sertularia spp. ( Roginskaya, 1987; Martynov British Isles (the type locality of this subspecies) & Korshunova, 2011), Hartlaubella gelatinosa and thereby confirm that it is conspecific with (Pallas, 1766), Laomedea flexuosa Alder, 1857 , our other specimens from different localities Obelia longissima (Pallas, 1766) , Sertularia across the North Atlantic. Present molecular argentea Linnaeus, 1758, Sertularia cupressina phylogenetic analysis (figs 1, 2) hereby confirms Linnaeus, 1758 (Alder & Hancock, 1845–1855; that C. concinna concinna has an amphiatlantic Swennen, 1961; Brown, 1980; Thompson & distribution, but North Pacific populations Brown, 1984; Picton & Morrow, 1994). A report belong to the separate subspecies C. concinna of the feeding on the athecate Tubularia bellatula subsp. nov. (see details below). The indivisa Linnaeus, 1758 (Hamond, 1972) needs range of C.concinna concinna is confirmed here a verification. from the White Sea to the Northeast Atlantic The maximum intragroup distance in C. coast, including occurrences in Eastern Canada, concinna concinna is 0.91%. The minimum COI the Netherlands, Norway, Russia, Sweden, the interspecies distance of9.28%is found between UK, and the Eastern USA. C. concinna Downloaded concinna from and Brill.com C. vasentsovichi 12/12/2023 04:16:22PM
via Open Access. This is an open access article distributed under the terms of the CC-BY 4.0 license.
https://creativecommons.org/licenses/by/4.0/
sp. nov. ( Table 1). Morphologically, the reported for this species by Williams & Gosliner nominative subspecies differs from C. concinna (1979). The reason the receptaculum in C. bellatula subsp. nov. by its larger maximum concinna s.l. (figs 19E, F) appears to be placed in body size (20 mm compared to 15 mm), and a a more proximal position than in the majority larger number of lateral denticles of the central of Cuthonella species (figs 19, 20) is possibly teeth of the radula. We also checked several because a channel from the receptaculum specimens of C. concinna s.l. from different to the female opening is partially concealed locations over large geographic distances, but within female gland mass. For a morphological were unable to confirm the presence of a narrow comparison with other Cuthonella species, see distal “bursa” (in addition to the receptaculum) table 2. Downloaded from Brill.com 12/12/2023 04:16:22PM via Open Access. This is an open access article distributed under the terms of the CC-BY 4.0 license. https://creativecommons.org/licenses/by/4.0/ Cuthonella concinna bellatula subsp. nov. sheen and a few scattered white or bluish ( Figs 1–3 View FIGURE 1 View FIGURE 2 View FIGURE 3 , 9 View FIGURE 9 , 19F View FIGURE 19 , 21 View FIGURE 21 ) spots. Ceratal tips with opaque white spots or ZooBank: http://urn:lsid:zoobank. bands. Rhinophores and oral tentacles semi-
o r g:a c t:8 2C F 5 3 9 3 - C F 7 0 - 4 2D F -A9 5A - transparent with white pigment on the tips E8869B4FBED8 (fig. 9A–F).
Cuthonella concinna s.l. auct., e.g. Jaws. Jaws broad, yellowish in colour. O’Donoghue, 1922: 162–163; Millen, 1983: 384; Masticatory processes of jaws covered with a Behrens, 1980: 98; Korshunova et al., 2018a: single row of distinct denticles (fig. 9G, H, I).
317, non Alder & Hancock, 1843 Radula. Radula formula about 17 × 0.1.0 Type material. Holotype. ZMMU Op-729, (specimen 8 mm in length), Central tooth
L = 6 mm length (live), NE Pacific, Rich elongate with protracted, pointed non- Passage, West of Pt. Glover, c. 12 m depth, compressedcusp(fig.9J,K,L). Centraltoothbears collected by Karin Fletcher, 13.07.2013. 3–5 lateral denticles. Cusp clearly delineated Paratypes. ZMMU Op-728, one spec., from the adjacent first lateral denticles .
L = 8 mm length (live), NE Pacific, Rich Reproductive system. Diaulic. Passage, Port Orchard, 10.7 m, collected by Hermaphroditic duct leads to a short, swollen Karin Fletcher, 18.04.2012. ZMMU Op-740, ampulla. Vas deferens short, without a distinct one spec., L = 7 mm length (live), NE Pacific, prostate. Supplementary gland short, inserts Rich Passage, West of Pt. Glover, c. 10 m depth, into the vas deferens a relatively long distance collected by Karin Fletcher, 02.02.2019. ZMMU from the elongated penial sheath. Penis Op-742, two spec., L = 5–6 mm length (live), conical, unarmed. Oviduct connects through NE Pacific, Rich Passage, West of Pt. Glover, the insemination duct into the female
c. 10 m depth, collected by Karin Fletcher, gland complex. Receptaculum seminis in a 30.04.2014. more proximal position than in most other Etymology. From Latin bellatula (“small Cuthonella species, and on a short stalk, with beautiful”) referring to the morphological only a weakly defined reservoir (fig. 19F). similarity, but smaller size, compared to Ecology. Lives on stones and rocks at the nominative subspecies C. concinna depths of c. 10–15 m, primarily found between concinna , for which the name “concinna ” March-September on its prey, the hydroids of has a similar meaning in Latin, “beautiful, the family Sertulariidae .
well-proportionated”. Distribution. Temperate NE Pacific waters External morphology. Body moderately from Alaska to Sonoma County, California .
narrow. Foot and tail moderate. Rounded foot Remarks. Morphologically C. concinna corners present. Oral tentacles moderate. bellatula subsp. nov. differs from the Rhinophores similar in size, or slightly longer nominative subspecies by its smaller than oral tentacles, smooth to slightly wrinkled. maximum body size (15 mm compared to Dorsal cerata elongate, arranged in continuous 20 mm), more varied ceratal colour (in C. rows.Potentially up to 6, but in present material concinna bellatula subsp. nov. the ceratal commonly up to 4 pre-anal unbranched ceratal colour ranges from pinkish to brownishrows. Anal opening acleioproctic. orange, reddish, grayish-purple to blackish, Colour (live). Background colour white to whereas in C. concinna concinna the colour yellowish white. Digestive gland diverticula is a rather uniform brownish to dark brown) pinkish, brownish-orange, reddish, grayish- and fewer number of lateral denticles on the purple to blackish. Cerata can have a bluish central teeth of the radula Downloaded from Brill.com (in C 12/12/2023. concinna 04:16:22PM via Open Access. This is an open access article distributed under the terms of the CC-BY 4.0 license.
https://creativecommons.org/licenses/by/4.0/
bellatula subsp. nov. the number of lateral California (CASIZ 181307a and CASIZ 181307d), denticles is commonly no more than 5, plus another specimen collected from the whereas in the nominative subspecies the same site on a different date (CASIZ 179469), number of denticles can reach up to 10). all putatively considered to be C. cocoachroma Cuthonella concinna bellatula and C. when they were collected, turned out to be cocoachroma are also likely to be mistaken for C. concinna (named C. concinna bellatula in each other. These two species are so similar this paper). These results were not discussed externally that two of the five specimens in details within the study which originally collected at the same time in Sonoma County, sequenced Downloaded the specimens from Brill(Cella.com 12 et/12al /2023., 2016 04:16):22PM via Open Access. This is an open access article distributed under the terms of the CC-BY 4.0 license. https://creativecommons.org/licenses/by/4.0/
but until then the southernmost limit of the C. concinna bellatula subsp. nov. from the range of C. concinna bellatula was Vancouver NE Pacific compared to the location of the Island, British Columbia, Canada. A record of nominative subspecies. In our opinion, the Cuthonella concinna s.l. from the Commander best decision in this case was the separation Islands ( Martynov , 1997) may also include of such forms as subspecies, which meets the a different species and needs further requirements of both the ICZN and general investigation. Several hydroid species were taxonomic practice when subspecies status previously reported as food objects for NE commonly implies a considerable geographic Pacific C. concinna s.l.: Bougainvillia glorietta separation. It is possible that with a really Torrey, 1904, Obelia commissuralis McCrady , large dataset this decision can be clarified, 1859 (currently also mentioned as Obelia but given the amount of currently available dichotoma (Linnaeus, 1758)), Coryne eximia data we prefer to consider it a subspecies.
Allman, 1859 ( Waters, 1966), and Sertularia sp. The maximum intragroup distance in C.
(Behrens & Hermosillo, 2005). These reports concinna bellatula is 1.67%. The minimum need verification. In the present study we COI intergroup distance of 9.74% is found found C. concinna bellatula subsp. nov. on between C. concinna bellatula subsp. nov.
hydroids of the family Sertulariidae . and C. vasentsovichi sp. nov. (table 1). For The maximum intragroup distance in C. a morphological comparison with other concinna bellatula subsp. nov. is 1.67%. At the Cuthonella species, see table 2.
same time, the minimum intergroup distance between C. concinna concinna and C. concinna Cuthonella denbei sp. nov.
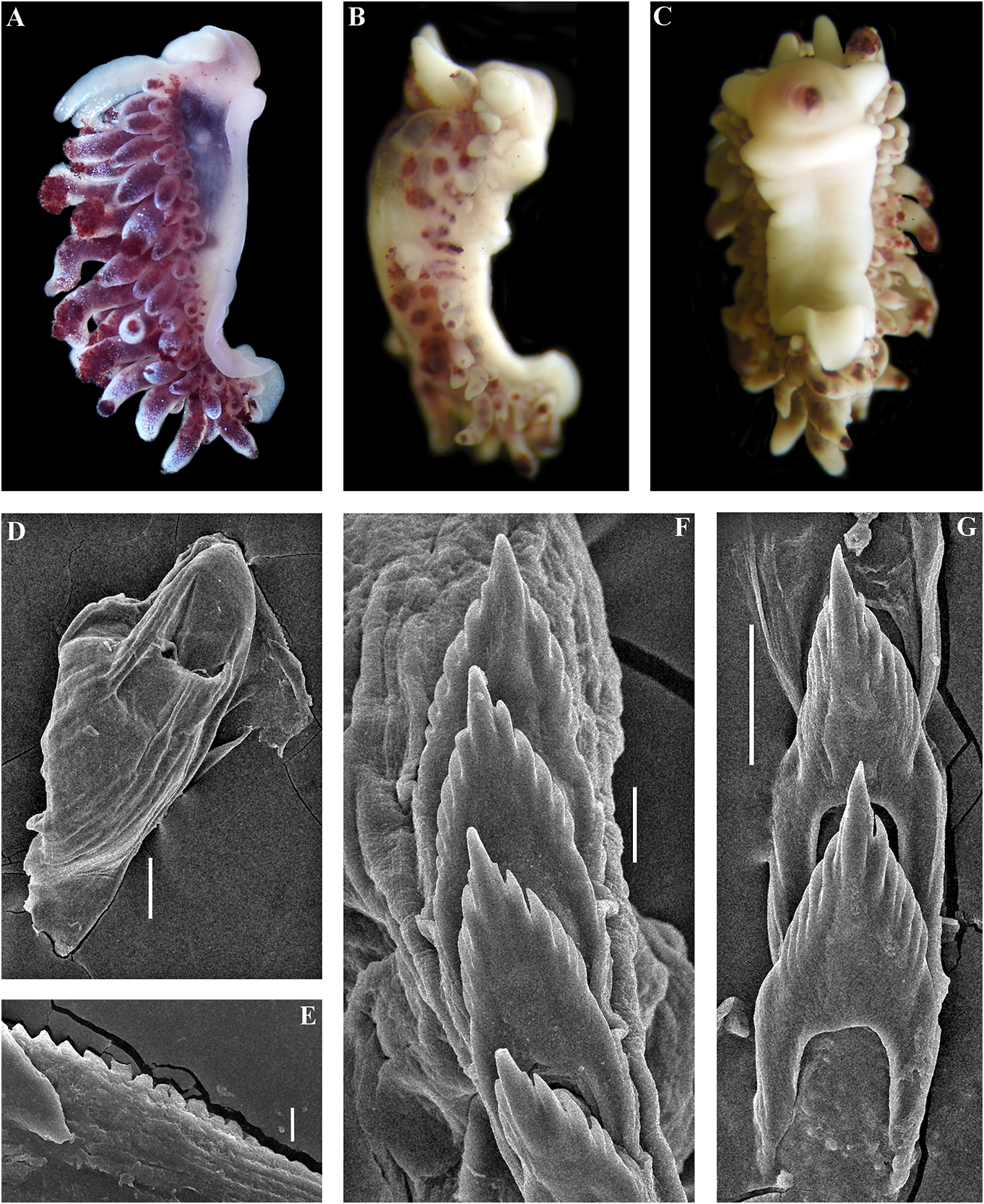
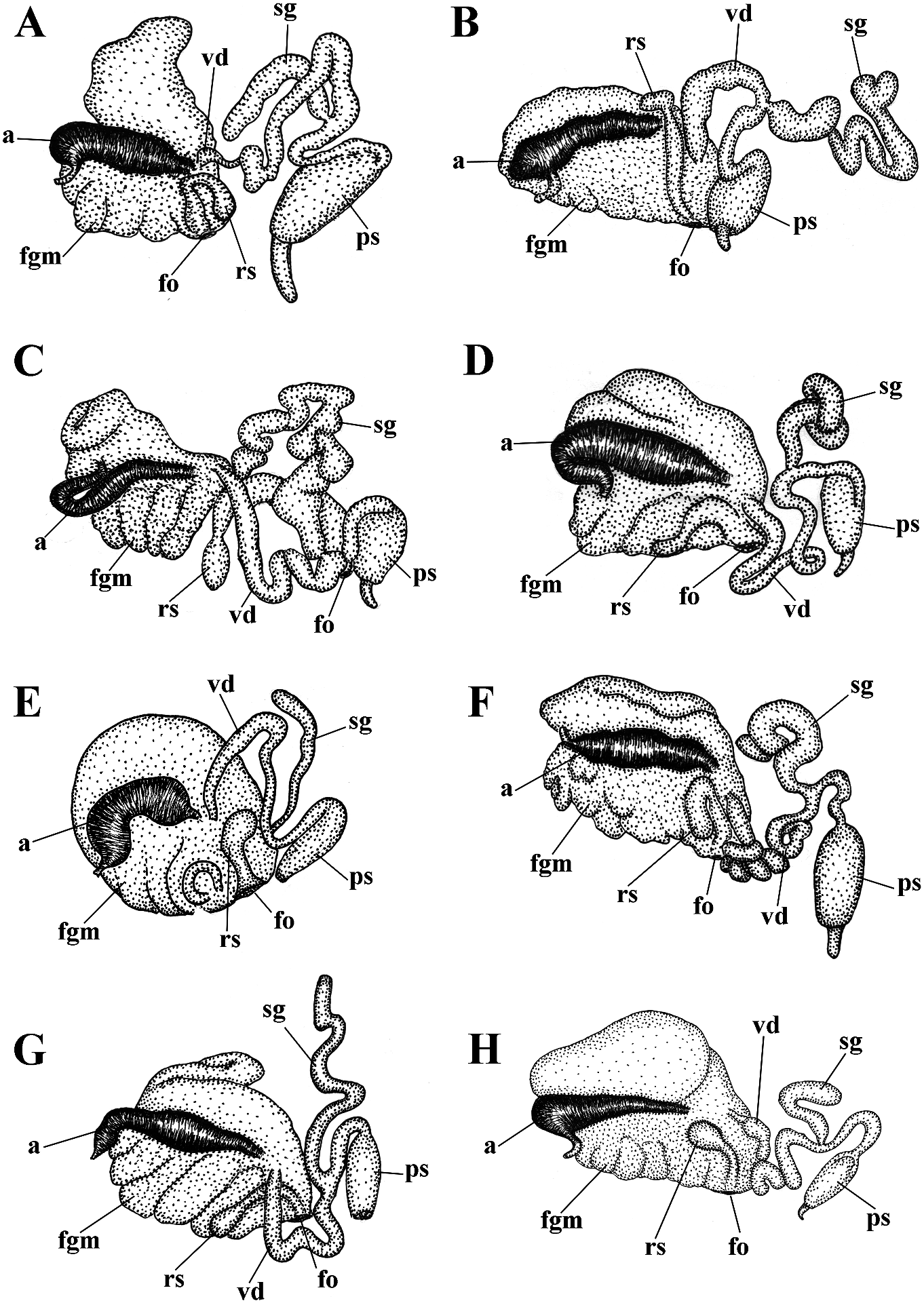
bellatula subsp. nov. is 1.07%. A considerable ( Figs 1 View FIGURE 1 , 2 View FIGURE 2 , 10 View FIGURE 10 , 20A View FIGURE 20 , 21 View FIGURE 21 )
intragroup genetic divergence suggests that ZooBank: http://urn:lsid:zoobank.org:act:9Dthe present North Pacific populations of C. D902C3-B8CA-4941-9712-11E77977DB77
concinna bellatula subsp. nov. may conceal Type material. Holotype. ZMMU Op-730, potentially more taxonomic diversity and may L = 6 mm length (preserved), Pacific Ocean, also have experienced a genetic introgression Middle Kuril Islands, Matua Island, Klyuv with Atlantic C. concinna concinna Cape , intertidal, collected by N.P. Sanamyan, occasionally introduced to the Pacific coast. 06.08.2017.
While making a decision on the taxonomic Paratypes. ZMMU Op-674, 1 spec., status of “ C. concinna ” from the NE Pacific L = 4.5 mm length (preserved), Pacific Ocean, we encountered a difficult situation because Middle Kuril Islands , Matua Island, Klyuv of the lack of a clear and universal definition Cape, intertidal, collected by N.P. Sanamyan ,
of a “species” concept. “ Cuthonella concinna ” 22.08.2016. ZMMU Op-673, 1 spec., L = 4 mm from the NE Pacific is case of a “species” (or length(preserved), Pacific Ocean,Middle Kuril a part of a group of species) which has likely Islands, Matua Island, Klyuv Cape, 16 m depth, only relatively recently started to diverge into on brown algae Thalassiophyllum clathrus ,
a separate species, where specimens already collected by N.P. Sanamyan, 23.08.2016. ZMMU show some morphological and molecular Op-741, 1 spec., L = 7 mm length (preserved), differences from C. concinna concinna , but Pacific Ocean , Middle Kuril Islands , Matua these differences are not distinct enough for a Island, Klyuv Cape, intertidal, collected by full species status. N.P. Sanamyan, 06.08.2017 .
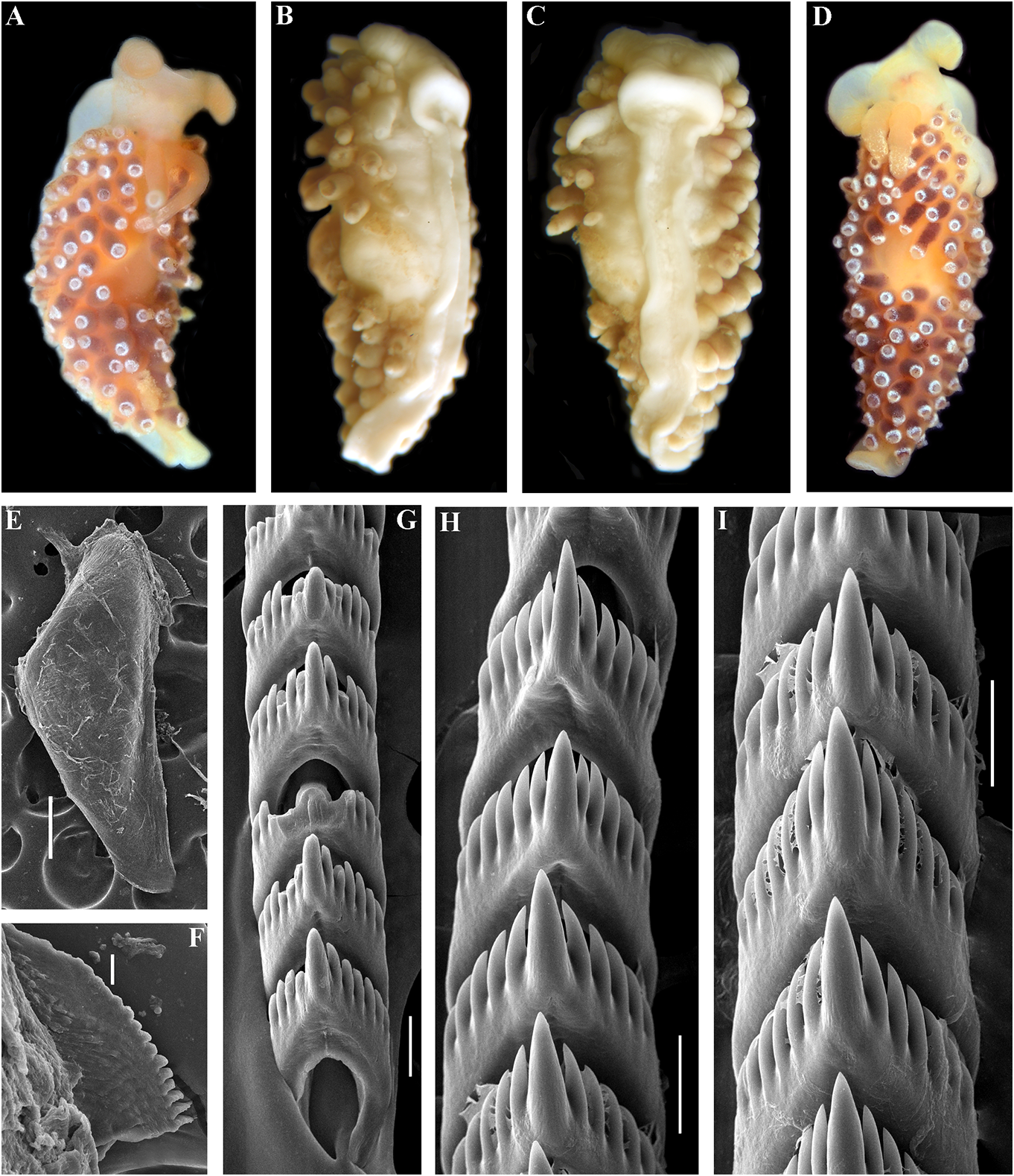
At the same time, a clear and significant Etymology. Named in honour of the geographic gap was revealed in the case of Japanese Downloaded merchant from clerk Brill.com Denbei12/12(/ Dembei 2023 04:16):22 – PM via Open Access. This is an open access article distributed under the terms of the CC-BY 4.0 license. https://creativecommons.org/licenses/by/4.0/ 伝兵Ñ(でん«え)(c. 1670–1714) from into the vas deferens a considerable distance Osaka who represented the first well- from the elongated penial sheath. Penis documented case of the appearance of a conical, unarmed. Oviduct connects through Japanese native in Russia. Denbei arrived in the insemination duct into the female gland Russia due to the shipwrecking of a Japanese complex. Receptaculum seminis in a distal vessel on the Kamchatka coast, followed by a position, on a short stalk, pear-shaped (fig. long drift through the Kuril Islands chain, the 20A). type locality of the present species.Afterwards, Ecology. Lives on soft bottoms with stones. Denbei had an audience with the Russian Intertidal to 16 m depth. tsar Peter the Great in 1702. This became an Distribution. Matua Island, Middle Kuril historic event since Denbei promoted the Islands. first official connections between Japan and Remarks. The maximum intragroup Russia, and also established the first school of distance in C. denbei sp. nov. is 0.30%. The Japanese language in Russia (e.g., Ikuta, 2008). minimum COI intergroup distance of 9.30% External morphology. Body moderately is found between C. denbei sp. nov. and C. narrow. Foot and tail broad. Oral tentacles concinna concinna (table 1). Morphological moderate. Rhinophores similar in size to oral analysis reveals considerable differences in tentacles, smooth to slightly wrinkled. Dorsal the denticulation of the central teeth between cerata elongate, thick, arranged in continuous C. denbei sp. nov. and related species C. rows. Up to 6 pre-anal unbranched ceratal benedykti , C. georgstelleri sp. nov., C. hiemalis rows. Anal opening acleioproctic. sp. nov., C. osyoro , and C. soboli (figs 7, 11, 12, Colour (live). Background colour yellowish 14, 17). For a morphological comparison with to more intense yellow-pinkish. Digestive other Cuthonella species, see table 2. gland diverticula within cerata brownish to dark-reddish. Ceratal tips with distinct ring- Cuthonella elenae Martynov, 2000 shaped opaque white pigment, sometimes Cuthonella elenae Martynov, 2000: 5–9 , Figs with few scattered white spots. Rhinophores 1–3; Martynov , 2006b: 290. light yellowish with a sparse white pigment at Remarks. Cuthonella elenae from the the top (fig. 10A, B). Chukchi Sea has been described in detail in Jaws. Jaws broad, yellowish in colour. Martynov (2000), which will not be repeated Masticatory processes of jaws covered with a here. This species has all the diagnostic single row of distinct denticles (fig. 10E, F). characters of the genus Cuthonella , including Radula. Radula formula 19 × 0.1.0 a supplementary gland inserted into the vas (specimen 4 mm in length). Central tooth deferens, which unambiguously places this elongated with a strongly protracted, pointed species in the genus Cuthonella , even in the non-compressed cusp (fig. 10G–I). Central absence of molecular data. At the same time, tooth bears up to 7 lateral denticles. Cusp this species evidently shows several unique clearly delineated from the adjacent first features, including small lobes around the lateral denticles. penial opening, a very broad penial sheath, Reproductive system. Diaulic. a relatively short supplementary gland, and Hermaphroditic duct leads to a relatively flattened oral tentacles. short, swollen ampulla. Vas deferens These characters clearly delineate this moderately long, without a distinct prostate. species from any other Cuthonella species, Supplementary gland relatively short, inserts including Downloaded the newly-described from Brill.com ones in 12/12/2023 04:16:22PM
via Open Access. This is an open access article distributed under the terms of the CC-BY 4.0 license.
https://creativecommons.org/licenses/by/4.0/
this study. By the presence of the many- ZooBank: http://urn:lsid:zoobank. branched digestive gland, this species shares o r g:a c t:F F 4 0 F D D4 - C F8 D -4 75 8- 8 9 9A -
a common plesiomorphic condition with 6D2A548E686B
the type species, C. abyssicola s.l. There are Type material. Holotype. ZMMU Op-670, no additional specimens collected since 1 spec., L = 14 mm length (preserved), Pacific the original description, and molecular Ocean , Middle Kuril Islands , Matua Island , phylogenetic data are not available for this Klyuv Cape , 16 m depth, collected by N.P. species. For a morphological comparison with Sanamyan, 28.08.2016.
other Cuthonella species, see table 2. Paratypes. ZMMU Op-672, 1 specimen ,
L = 12 mm length (preserved), same locality, Cuthonella georgstelleri sp. nov. date and collector as holotype. ZMMU Op-731 ,
( Figs 1 View FIGURE 1 , 2 View FIGURE 2 , 11 View FIGURE 11 , 20B View FIGURE 20 , 21 View FIGURE 21 ) 1 spec., L Downloaded = 10 mm from length Brill.com (preserved 12/12/2023), Pacific 04:16:22PM via Open Access. This is an open access article distributed under the terms of the CC-BY 4.0 license. https://creativecommons.org/licenses/by/4.0/ Ocean, Middle Kuril Islands, Matua Island, made substantial efforts to study the North Klyuv Cape, 14 m depth, collected by N.P. Pacific region, and particularly the Kuril Sanamyan, 20.08.2017. and Commander Islands, but met a very Etymology. The name is in honour of the unfortunate fate. Georg Steller was the only prominent 18th-century scientist Georg professional scientist who saw and personally Wilhelm Steller (1709–1746) of German described the now extinct Steller’s sea cow origin who served in the Russian Empire and ( Hydrodamalis Downloaded gigas from). Georg Brill.com Steller 12/12/2023 is also 04:16:22PM
via Open Access. This is an open access article distributed under the terms of the CC-BY 4.0 license.
https://creativecommons.org/licenses/by/4.0/ considered the first European who stepped the insemination duct into the female gland onto the land of Alaska ( Stejneger, 1936). complex. Receptaculum seminis distal, on a In 1742, Georg Steller was falsely accused of long stalk, compressed (fig. 20B). organizing a rebellion against the Russian Ecology. Lives on soft bottoms with stones Government in Kamchatka and was forced at depths of 14– 16 m. to return to mainland Russia, but on the way Distribution. Matua Island, Middle Kuril to Moscow he died in the Siberian area of Islands. Tyumen. The majority of the descriptions of Remarks. The maximum intragroup his collected materials and prepared notes (in distance in C. georgstelleri sp. nov. is 0.30%. his diaries) was published after his death. The minimum COI intergroup distance External morphology. Body moderately of 7.75% is found between C. georgstelleri wide. Foot and tail broad. Oral tentacles sp. nov. and C. benedykti sp. nov. (table 1). moderate. Rhinophores similar in size to Phylogenetically, C. georgstelleri sp. nov. is oral tentacles, smooth to slightly wrinkled. close to C. soboli , C. osyoro , C. ainu sp. nov., Dorsal papillae elongate, thick, arranged in and C. benedykti sp. nov. Morphological continuous rows. Up to8 pre-anal unbranched analysis reveals considerable differences in or partially branched ceratal rows. Anal the denticulation of the central teeth between opening cleioproctic. C. georgstelleri and the related species C. ainu Colour (live). Background colour whitish. sp. nov., C. hiemalis , C. osyoro and C. soboli (figs Digestive gland diverticula light orange- 6, 12, 14, 17). For a morphological comparison brownish and salmon-brownish to dark, with other Cuthonella species, see table 2. intensive brownish reddish. Ceratal tips with extended opaque white band or spot, Cuthonella hiemalis ( Roginskaya, 1987) sometimes with few white small spots. ( Figs 1 View FIGURE 1 , 2 View FIGURE 2 , 12 View FIGURE 12 , 20C View FIGURE 20 , 21 View FIGURE 21 ) Rhinophores whitish to light yellowish with Cuthona hiemalis Roginskaya, 1987:197–199 ; slight disperse white pigment (fig. 11A–C). Fig. 118 was supposed to illustrate this species Jaws. Jaws broad, yellowish in colour. in the original description, but it was omitted Masticatory processes of jaws covered with a in the publication due to an editorial error. single row of denticles (fig. 11E, F). Cuthonella hiemalis ( Roginskaya, 1987) – Radula. Radula formula 30 × 0.1.0 Martynov , 1992: 23; Martynov , 2006b: 290; (specimen 14 mm in length). Central tooth Martynov & Korshunova, 2011: 195–197, figs. elongated with a strongly protracted, pointed Type material. The previously indicated non-compressed cusp (fig. 11 G, H). Central ( Martynov , 2006b) syntypes series (including tooth bears up to 10 lateral denticles. Cusp labelled, but not published by Roginskaya clearly delineated from the adjacent first (1987) “ holotype ”) is not traceable. A syntype lateral denticles. wasdiscovered,accompaniedbyalabelwritten Reproductive system. Diaulic. by I.S. Roginskaya. The syntype was collected Hermaphroditic duct leads to a relatively over the same period of time in the same short, swollen ampulla. Vas deferens locality with the previously mentioned type moderate in length, without a distinct series: ZMMU Op-732, L = 14 mm (preserved), prostate. Supplementary gland long, inserts White Sea, Rugozerskaya Bay, intertidal, into the vas deferens at a considerable length collected by I.S. Roginskaya, 7.02.1962. from the elongated penial sheath. Penis Other materials. ZMMU Op-186, eggconical, unarmed. Oviduct connects through mass and Downloaded juveniles from (preserved Brill.com), White 12/12/2023 Sea, 04:16:22PM
via Open Access. This is an open access article distributed under the terms of the CC-BY 4.0 license.
https://creativecommons.org/licenses/by/4.0/ Rugozerskaya Bay , intertidal, collected by T. A. Dorsal cerata elongate, thick, arranged in Korshunova , and A. V. Martynov , 14.06.2009. continuous rows. Up to 7 pre-anal unbranched ZMMU Op-666, 5 spec., L = c. 5 mm each ceratal rows. Anal opening acleioproctic. (preserved), White Sea, Rugozerskaya Bay, Colour (live). Background colour yellowish, collected by A.E. Zhadan, 2009–2010. sometimes with dispersed orange pigment. External morphology. Body moderately wide. Digestive gland diverticula in cerata from
Foot and tail broad. Oral tentacles moderate. dark beige to dark brown. Ceratal tips with Rhinophores similar in size or slightly shorter peculiar opaque bluish to whitish subapical than oral tentacles, smooth to slightly wrinkled. band or spot Downloaded, without from opaque Brill.com white12/12pigment /2023 04:16.:22PM via Open Access. This is an open access article distributed under the terms of the CC-BY 4.0 license. https://creativecommons.org/licenses/by/4.0/ Rhinophores yellowish without distinct white osyoro , and C. soboli Martynov, 1992 (figs pigment at the top (fig. 12A). 7, 10, 11, 14, 17). Reproduction occurs in the Jaws. Jaws broad, yellowish in colour. wintertime. Cuthonella hiemalis feeds on the Masticatory processes of jaws covered with a athecate hydroids Clava multicornis (Forsskål, single row of distinct denticles (fig. 12D, E). 1775), Sarsia lovenii (M. Sars, 1846) , and on Radula. Radula formula 23 × 0.1.0 the thecate Dynamena pumila (Linnaeus, (specimen 14 mm in length). Central tooth 1758) ( Roginskaya, 1987). For a morphological elongated with a strongly protracted, pointed comparison with other Cuthonella species, see non-compressed cusp (fig. 12F, G). Central table 2. tooth bears up to 12 lateral denticles. Cusp clearly delineated from the adjacent first Cuthonella orientosiberica sp. nov. lateral denticles. ( Figs 1 View FIGURE 1 , 2 View FIGURE 2 , 13 View FIGURE 13 , 20D View FIGURE 20 , 21 View FIGURE 21 ) Reproductive system. Diaulic. ZooBank: http://urn:lsid:zoobank.org:act:- Hermaphroditic duct leads to a relatively long, 106FE0CB-1D90-4EDC-B649-BB4AE07932C7 folded, non-swollen ampulla.Vas deferens long, Type material. Holotype. ZMMU Op-184, without a distinct prostate. Supplementary L = 3.5 mm length (preserved), East Siberian gland long, inserts into the vas deferens a Sea, 70° 09.90´N, 161° 16.18´E, depth 9 m, considerable distance from the elongated collected by A.Y. Voronkov, 06.09.2004. penial sheath. Penis conical, unarmed. Oviduct Paratypes. ZMMU Op-733, L = 4 mm connects through the insemination duct into length (preserved), East Siberian Sea, 70° the female gland complex. Receptaculum 09.90´N, 161° 16.18´E, depth 9 m, collected seminis in a distal position, on a long massive by A.Y. Voronkov, 06.09.2004. ZMMU Op-185, stalk, oval (fig. 20C). L = 4 mm length, East Siberian Sea, 69° 10.50´Ecology. Lives on stones and rocks. N, 169° 52.87´E, depth 15 m, collected by A.Y. Intertidal to 5 m depth. Larvae exhibit direct Voronkov, 07.09.2004. development without a planktonic stage. Etymology. The name refers to the type Distribution. Currently known only from locality. Oriento-, Lat. “belonging to the east” the White Sea, possibly it also occurs in the and the Siberian Sea. Barents Sea. External morphology. Body moderately Remarks. According to the molecular wide. Foot and tail broad. Oral tentacles phylogenetic analysis, Cuthonella hiemalis moderate. Rhinophores similar in size to oral forms a separate clade sister to the clade tentacles, smooth to slightly wrinkled. Dorsal containing C. soboli , C. osyoro , C. benedykti cerata elongate, thick, arranged in continuous sp. nov., C. georgstelleri sp. nov., and C. ainu rows. Up to 6 pre-anal unbranched ceratal sp. nov. (figs 1, 2). The maximum intragroup rows. Anal opening acleioproctic. distance in C. hiemalis is 0.31%. The Colour (live). Background colour yellowish minimum COI intergroup distance of 8.37% white. Digestive gland diverticula light is found between C. hiemalis and C. osyoro brownish to yellowish in juveniles and (table 1). brownish to dark brown in adults. Ceratal Morphological analysis reveals tips whitish. Rhinophores light yellowish with considerable differences in the denticulation white pigment at the top. of the central teeth between C. hiemalis and Jaws. Jaws broad, yellowish in colour. the related species C. benedykti sp. nov., C. Masticatory processes of jaws covered with a denbei sp. nov., C. georgstelleri sp. nov., C. single row Downloaded of denticles from (fig Brill.com. 13D, E). 12/12/2023 04:16:22PM
via Open Access. This is an open access article distributed under the terms of the CC-BY 4.0 license.
https://creativecommons.org/licenses/by/4.0/
Radula. Radula formula 28 × 0.1.0 (specimen cusp (fig. 13F, G). Central tooth bears up to 9
4 mm in length). Central tooth elongated with lateral denticles. Cusp clearly delineated from a strongly protracted, pointed non-compressed the adjacent Downloaded first lateral from denticles Brill.com. 12/12/2023 04:16:22PM via Open Access. This is an open access article distributed under the terms of the CC-BY 4.0 license. https://creativecommons.org/licenses/by/4.0/
Reproductive system. Diaulic. Herma- stones, collected by T.A. Korshunova and phroditic duct leads to a relatively long, A.V. Martynov , 16.09.2014. ZMMU Op-607, 1 folded, swollen ampulla. Vas deferens spec., L = 4 mm length (preserved), the Sea moderately long, without a distinct prostate. of Japan, Spaseniya Bay, 0.3–0.7 m, stones, Supplementary gland long, inserts into the collected by T.A. Korshunova and A.V. vas deferens a considerable distance from Martynov , 16.09.2014. ZMMU Op-524, 1 spec., the elongated penial sheath. Penis conical, L = 3 mm length (preserved), the Sea of Japan, unarmed. Oviduct connects through the Gornostay Bay, 0.5–0.7 m, stones, collected insemination duct into the female gland by T.A. Korshunova and A.V. Martynov , complex. Receptaculum seminis in a distal 18.09.2014. ZMMU Op-604, 1 spec., L = 4 mm position, on a short stalk, oval (fig. 20D). length (live), the Sea of Japan, Spokoinaya Ecology. Lives on stones and rocks at depths Bay, 3–5 m, brown algae, stones, collected of c. 9– 15 m. by T.A. Korshunova and A.V. Martynov , Distribution. Arctic Seas – East Siberian 25.09.2014. ZMMU Op-613, 1 spec., L = 9 mm Sea, Laptev Sea, possibly also Chukchi Sea. length (live), the Sea of Japan, Spokoinaya Bay, Remarks. According to the morphological 3–5 m, brown algae, stones, collected by T.A. data, Cuthonella orientosiberica sp. nov. is Korshunova and A.V. Martynov , 25.09.2014. similar to C. hiemalis ( Roginskaya, 1987) and External morphology. Body moderately C. soboli Martynov, 1992 . However, there are wide. Foot and tail broad. Oral tentacles differences in the denticulation of the central moderate. Rhinophores similar in size to teeth between C. orientosiberica sp. nov. and oral tentacles, smooth to slightly wrinkled. the species C. hiemalis and C. soboli (figs 12, Dorsal papillae elongate, thick, arranged in 17). The available material was not suitable for continuous rows. Up to 6 pre-anal unbranched molecular study, despite several attempts to ceratal rows. Anal opening acleioproctic. extract DNA. For a morphological comparison Colour (live). Background colour transwith other Cuthonella species, see table 2. lucent whitish, sometimes slightly yellowish.
Digestive gland diverticula brownish to dark Cuthonella osyoro (Baba, 1940) red or violet. Ceratal tips with variously
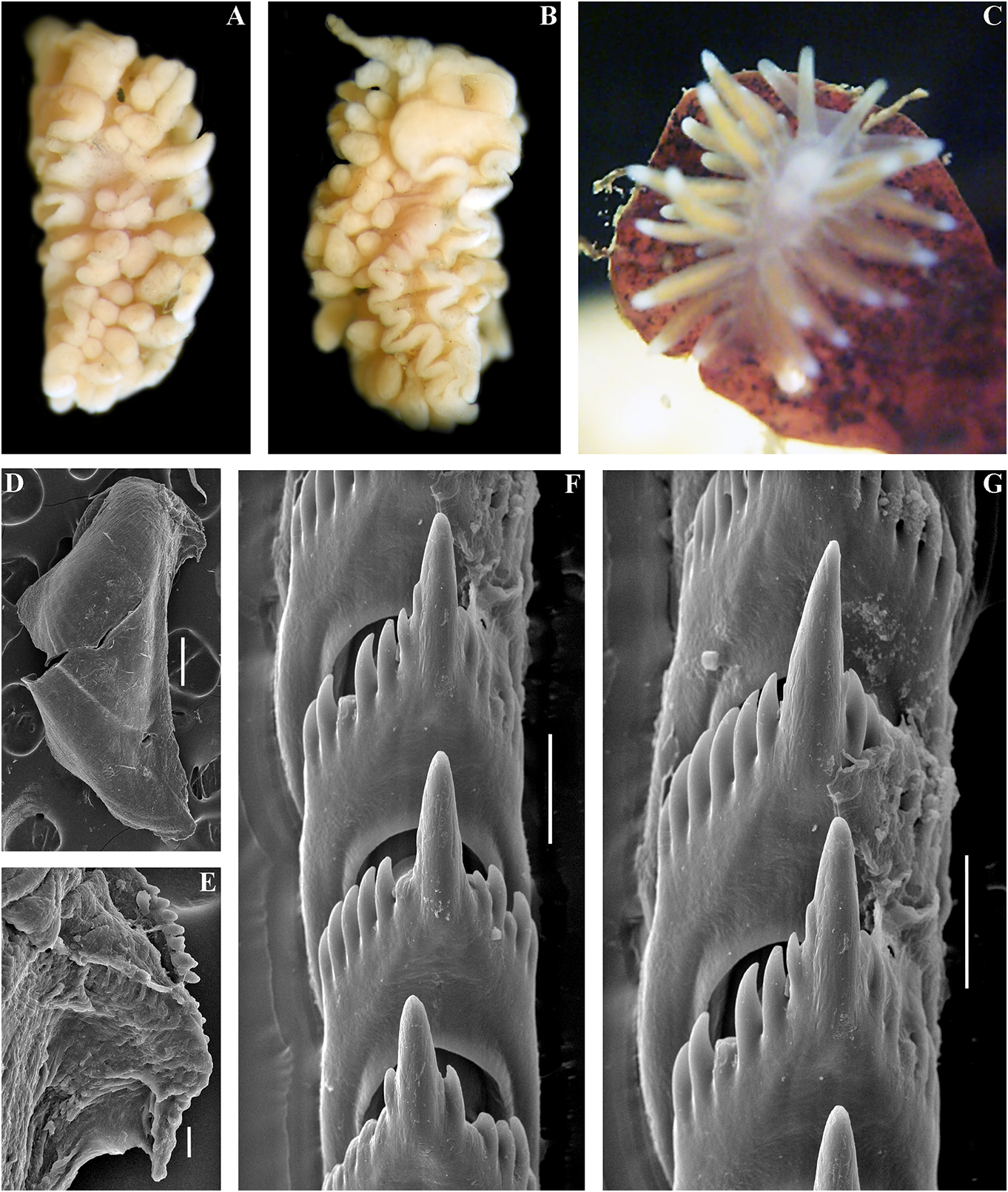
( Figs 1 View FIGURE 1 , 2 View FIGURE 2 , 14 View FIGURE 14 , 20E View FIGURE 20 , 21 View FIGURE 21 ) expressed red or orange band or spot. On Cuthona osyoro Baba, 1940: 108 , fig. 8 cerata white streaks or dots may be present. Cuthonella osyoro (Baba, 1940) – Martynov , Rhinophores semi-transparent, sometimes 2006b: 290; Martynov , Korshunova, 2011, with white pigment at the top (fig. 14A–G). partim, only the fig. on page 198. Jaws. Jaws broad, yellowish in colour. Type material. Holotype, lost. Neotype Masticatory processes of jaws covered with a ZMMU Op-743, L = 4 mm length (live), the Sea single row of distinct denticles (fig. 14H, I). of Japan, Spokoinaya Bay, 3–5 m depth, brown Radula. Radula formula 26× 0.1.0 (specimen algae, stones, collected by T.A. Korshunova 9 mm in length), 24 × 0.1.0 (specimen 8 mm and A.V. Martynov , 25.09.2014. in length). Central tooth elongated with a Other material. ZMMU Op-734, L = c. strongly protracted, pointed non-compressed 15 mm length (preserved, c. 25 mm live), the cusp (fig. 14 J–M). Central tooth bears up to 11 Sea of Japan, Vostok Bay, 0.2–0.4 m, stones, lateral denticles. Cusp clearly delineated from collected by A.V. Martynov , 18.02.1990. ZMMU the adjacent first lateral denticles. Op-606, 1 spec., L = 3 mm length (preserved), Reproductive system. Diaulic. Hermathe Sea of Japan, Spaseniya Bay, 0.3–0.7 m, phroditic Downloaded duct leads from to Brill.com a relatively short, 12/12/2023 04:16:22PM
via Open Access. This is an open access article distributed under the terms of the CC-BY 4.0 license.
https://creativecommons.org/licenses/by/4.0/ swollen, convoluted ampulla. Vas deferens painting by Baba (fig. 14G, M) to confirm the moderately long, without a distinct prostate. identity of this species.In the summertime in the Supplementary gland long, inserts into the intertidal and upper subtidal zones of the Sea of vas deferens a considerable distance from the Japan juveniles of C. osyoro are quite common, elongated penial sheath. Penis conical, unarmed. and therefore Baba (1940) described a juvenile Oviduct connects through the insemination duct of exactly this species from a neighbouring into the female gland complex. Receptaculum location at the Hokkaido coast of the Sea of seminis in a distal position, on a short stalk, Japan. In order to avoid confusion between two rather rounded (fig. 20E). sympatric sister species C. soboli and C. osyoro , Ecology. Lives on stones and rocks. neotype is selected here for C. osyoro (fig. 14A). Intertidal to 20 m depth. The neotype is essentially similar in colouration Distribution. The Northern part of the Sea and size to the first description of C. osyoro and of Japan, Hokkaido, possibly also the Southern was collected in the Sea of Japan across from Kuril Islands. the type locality in Hokkaido. The food objects Remarks. According to the molecular of C. osyoro possibly include athecate hydroids. phylogenetic analysis, Cuthonella osyoro forms For a morphological comparison with other a sister clade with C.soboli and is also a member Cuthonella species, see table 2. of a larger clade which includes C. hiemalis , C. benedykti sp. nov., C. georgstelleri sp. nov., Cuthonella punicea (Millen, 1986) and C. ainu sp. nov. (figs. 1, 2). The maximum ( Figs 1 View FIGURE 1 , 2 View FIGURE 2 , 15 View FIGURE 15 , 20F View FIGURE 20 , 21 View FIGURE 21 ) intragroup distance in C. osyoro is 1.15%. The Cuthona punicea Millen, 1986: 1356–1358 , minimum COI intergroup distance of 2.89% is figs 1–7. found between C. osyoro and C. soboli (table Cuthonella punicea (Millen, 1986) – 1). Cuthonella osyoro differs externally from Martynov , 1992: 23. the closely related species C. soboli (fig. 17) in Non “ Tenellia” pustulata sensu Cella always having red or orange spots or rings on et al., 2016 (= Zelentia nepunicea , see the ceratal tips. This variable-sized reddish or discussion in Korshunova et al., 2017a, 2018b; orange spot at the ceratal tips is present in all misidentification of Zelentia nepunicea with specimens of C. osyoro that we have observed, Cuthonella punicea in Cella et al., 2016). even in small juveniles, which was confirmed Type material. Holotype. CASIZ 060327, NE in the present study (fig. 14E, F). However, Pacific, Canada, British Columbia, Plumper prior to this study C. osyoro specimens were Islands, 50° 36´N, 126° 49´W, 13 m depth, sometimes mistaken for C. soboli because the rocks, collected by S.V. Millen, 06.05.1985. pale orange ceratal spot on juvenile C. osyoro Topotypes. ZMMU Op-735, 3 specimens, specimens that were sequenced (fig. 14D, L = 7 mm, 7.2 mm,and 4 mm length(preserved), for example) was inadvertently overlooked NE Pacific, Canada, British Columbia, Plumper (Korshunova et al., 2017b, 2018a). Islands, 50° 36´N, 126° 49´W, 20–22 m, rocks, In the original description, Baba (1940) collected by S.V. Millen, 07.05.1983. KM 765, described a juvenile specimen from Hokkaido 1 specimen, L = 21 mm, NE Pacific, Canada, which is identifiable by the presence of red British Columbia, Plumper Islands, 8.2–17.7 markings on the ceratal apices and patterns of m, collected by Karin Fletcher, 12.04.2019. KM the radula. In the present study, we specially 766, L = 15 mm, NE Pacific, Canada, British visited the archive of Baba’s material in Osaka Columbia, Plumper Islands, 8.2–17.7 m, and have presented a copy of the original collected Downloaded by Karin from Fletcher Brill.com, 12.04.2019. 12/12/2023 04:16:22PM
via Open Access. This is an open access article distributed under the terms of the CC-BY 4.0 license.
https://creativecommons.org/licenses/by/4.0/
External morphology. Body wider in front gland diverticula from purple to dark wine-
than behind. Foot and tail broad. Oral tentacles red,(pomegranate colour). Cerata with moderate. Rhinophores smooth and similar in white spots or streaks on surface. Cerata size or slightly longer than the oral tentacles. with dispersed white tips. Rhinophores Dorsal papillae elongate,arranged in continuous and oral tentacles semi-transparent to light rows.Up to 8 pre-anal partially branched ceratal yellowish and often frosted with varying rows. Anal opening cleioproctic. amounts of white pigment for almost the Colour (live). Background colour pinkish- entire length, or the pigment can form a yellowish white to pale peach. Digestive line (fig. 15Downloaded A–C). from Brill.com 12/12/2023 04:16:22PM via Open Access. This is an open access article distributed under the terms of the CC-BY 4.0 license. https://creativecommons.org/licenses/by/4.0/ Jaws. Jaws broad, yellowish in colour. Sandra Millen in 1983 (fig. 15D–F, I, J, M, N), Masticatory processes of jaws covered with a and confirm their essential similarity with single row of distinct denticles (fig. 15G–J). the recent specimens collected from the type Radula. Radula formula 12 × 0.1.0 (specimen locality (fig. 15A–C). 7 mm in length), c. 18 × 0.1.0 (spec. 15 mm The profound morphological differences in length). Central tooth elongated with a between this cuthonellid, Cuthonella punicea , strongly protracted, pointed non-compressed and the trinchesiid Zelentia nepunicea have cusp (fig. 15K–N). Central tooth bears up to 7 previously been highlighted(see Korshunova et lateral denticles. Cusp clearly delineated from al., 2017a, 2018b). Those differences combined the adjacent first lateral denticles. with the examination of original topotypes Reproductive system. Diaulic. and fresh material for molecular analysis in Hermaphroditic duct leads to relatively short, this study provide irrefutable evidence that straight, swollen ampulla. Vas deferens long, the synonymy of those two species proposed without a distinct prostate. Supplementary by Cella et al. (2016) is not valid. gland long, inserts into the vas deferens a A cuthonellid from the NW Pacific (Matua considerable distance from the elongated Island), Cuthonella sandrae sp. nov., with a penial sheath. Penis conical, narrow, unarmed. similar purple colour is a sister species to C. Oviduct connects through the insemination punicea (figs 1, 2), but differs significantly duct into the female gland complex. Tubular according to genetic distances, molecular receptaculum seminis in a distal position, phylogenetic analysis, external patterns of the without a distinct reservoir (fig. 20F). digestive gland branches and the shape of the Ecology. Lives on soft bottoms with stones radular teeth (figs 15, 16). at depths of 8–33 m, on or near its hydroid Cuthonella punicea forms the clade sister prey, the large athecate species Zyzzyzus to the clade containing Cuthonella sandrae rubusidaeus Brinckmann-Voss & Calder , sp. nov. and C. abyssicola kryos subsp. nov. 2013 (Millen, 1986; Brinckmann-Voss & (fig. 1). The maximum intragroup distance Calder, 2013). Eggs hatch after 14–15 days as in C. punicea is 0.76%. The minimum COI planktotrophic veligers. intergroup distance of 6.85% is found between Distribution. Northeast Pacific, so far only C. punicea and C. sandrae sp. nov (table 1). known from the Plumper Island group located For a morphological comparison with other on the northern end of Vancouver Island, Cuthonella species, see table 2. British Columbia. Remarks. In the present study, molecular Cuthonella sandrae sp. nov. data from recently collected Cuthonella ( Figs 1 View FIGURE 1 , 2 View FIGURE 2 , 16 View FIGURE 16 , 21 View FIGURE 21 ) punicea from the exact type locality of the ZooBank: http://urn:lsid:zoobank.org:act:- Plumper Islands ( Canada, British Columbia) F04A4DCF-8A29-45BA-88B7-9749DA6E4C72 is presented for the first time (figs 1, 2, 21). Type material. Holotype. ZMMU Op-671, According to that molecular phylogenetic 1 spec., L = 4 mm length (preserved), Pacific data, the species originally described by Ocean, Middle Kuril Islands, Matua Island, Millen (1986) as “ Cuthona ” belongs to the Klyuv Cape, 14 m depth, collected by N.P. same subclade with the type species of Sanamyan, 20.08.2017. Cuthonella , C. abyssicola , and to a larger clade Etymology. Named in honour of Sandra with all other Cuthonella (figs 1, 2). We also Violet Millen, a prominent nudibranch examined topotypes originally collected by researcher Downloaded of North from Pacific Brill.com nudibranch 12/12/2023 04:16:22PM
via Open Access. This is an open access article distributed under the terms of the CC-BY 4.0 license.
https://creativecommons.org/licenses/by/4.0/ molluscs. Most remarkably, in the present continuous rows. Up to c. 7 pre-anal branched study we discovered a new species from ceratal rows. Anal opening cleioproctic.
the Kuril Islands which is a sister species to Colour (live). Background colour semi- Cuthonella punicea (Millen, 1986) previously transparent white with purple-bluish areas. described by Sandra Millen from British Digestive gland diverticula in the cerata Columbia, and which is similar to the new violet-purple. Ceratal tips with dispersed species in its reddish-purple colouration white pigment and bluish hue. Rhinophores
(fig. 15A–C) but with substantial genetic and semi-transparent with sparse white pigment radular differences. at the top (fig. 16A).
External morphology. Body moderately Jaws. Jaws broad, yellowish in colour. wide. Foot and tail broad. Oral tentacles Masticatory processes of jaws covered with a moderate. Rhinophores similar in size to single row of distinct denticles (fig. 16D, E).
oral tentacles, smooth to slightly wrinkled. Radula. Radula formula 12 × 0.1.0 (specimen Dorsal papillae elongate, thick, arranged in 4 mm in length Downloaded). Central from Brill.com tooth12/12 elongated /2023 04:16:22PM via Open Access. This is an open access article distributed under the terms of the CC-BY 4.0 license. https://creativecommons.org/licenses/by/4.0/
with a strongly protracted, pointed non-compressed cusp (fig. 16F, G). Central tooth bears up to 8 lateral denticles. Cusp relatively unclearly delineated from the adjacent first lateral denticles.
Reproductive system is underdeveloped in the holotype and not suitable for a detailed study.
Ecology. Lives on soft bottoms with stones at 14 m depth.
Distribution. Matua Island, Middle Kuril Islands.
Remarks. According to the molecular phylogenetic analysis, Cuthonella sandrae sp. nov. is closest to C. punicea (Millen, 1986) and C. abyssicola kryos (figs 1, 2). The minimum COI intergroup distance of 6.85% is found between C. sandrae sp. nov. and C. punicea ( Table 1). Morphological analysis and molecular analysis reveal considerable differences between C. sandrae sp. nov. and any other species of the genus Cuthonella , including the externally similar C. punicea (fig. 15). Particularly in C. sandrae sp. nov. a continuous series of small branches of the digestive gland forms between the anterior and posterior parts of the digestive gland even at a relatively small size. In contrast, C. punicea has a rather linear, only weakly branched digestive gland which does not form a continuous row in between the anterior and posterior parts of the digestive gland even at a considerably larger size. The radular patterns between C. punicea and C. sandrae sp. nov. are also considerably different (figs 15M, N and 16F, G). For a morphological comparison with other Cuthonella species, see table 2.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |