
Eulimnadia gnammaphila, Brian V Timms, 2016
|
publication ID |
https://doi.org/10.11646/zootaxa.4066.4.1 |
|
publication LSID |
lsid:zoobank.org:pub:B0F56A57-C033-48C6-BB66-B007A93FC259 |
|
DOI |
https://doi.org/10.5281/zenodo.4685678 |
|
persistent identifier |
https://treatment.plazi.org/id/038C2831-6532-A13D-94B3-F98EE0B0AE3C |
|
treatment provided by |
Plazi |
|
scientific name |
Eulimnadia gnammaphila |
| status |
sp. nov. |
Eulimnadia gnammaphila View in CoL sp. nov.
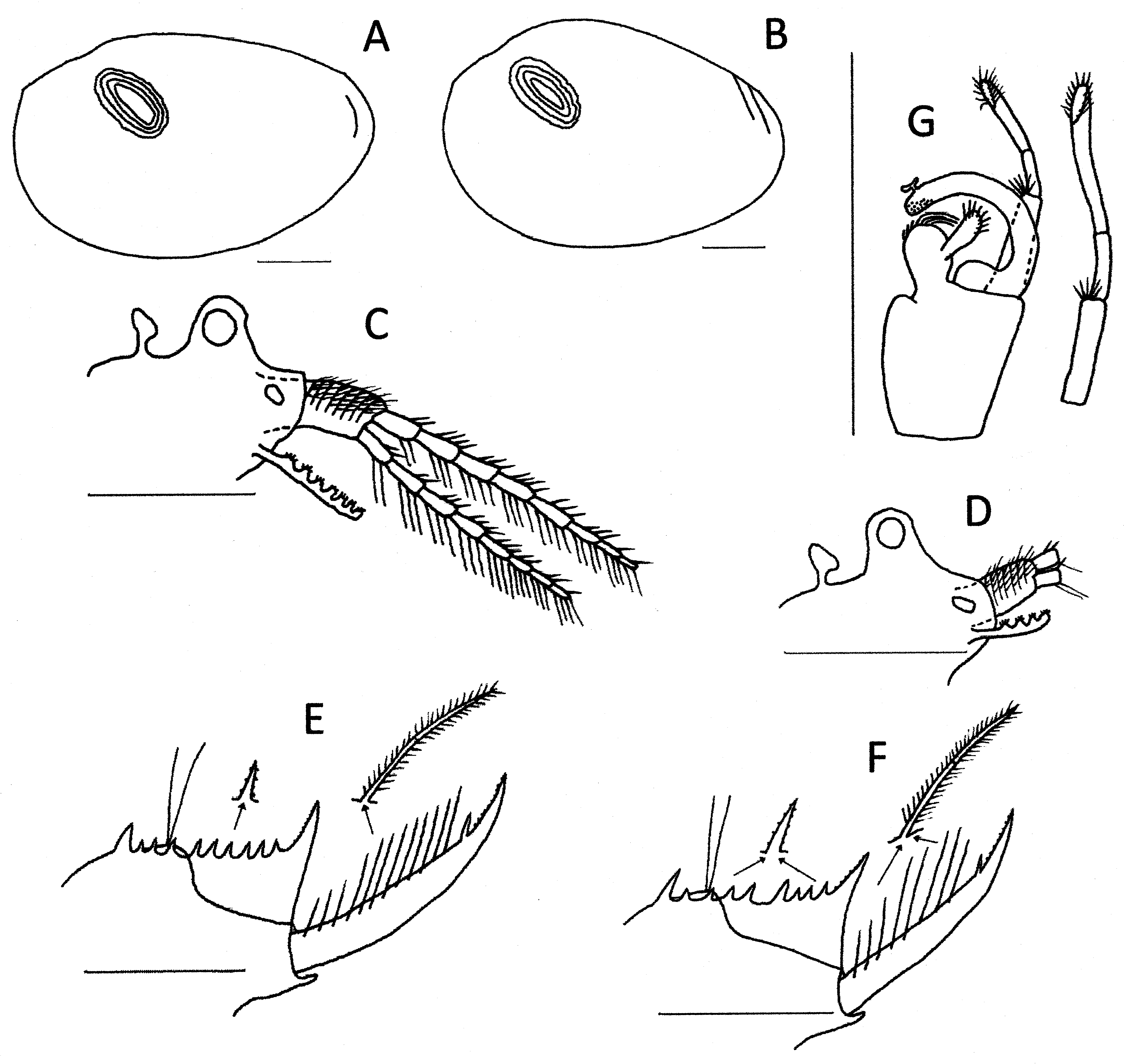
( Figs. 9 View FIGURE 9 B,C,D; 10,11,12)
Etymology. The species is named for the habitat from which all specimens have been taken across southern Australia. Gnammas are rock holes usually in granite, the term being derived from the Nyungar aboriginal language of south-west of Western Australia, while ‘phila’ is derived from the Greek ‘philos’ indicating love for something.
Type locality. South Australia, Peela Rocks, 25.5 km ENE of Wudinna, pit gnamma #3 (sensu Timms and Rankin, 2015), 33o 00’ 09.6”S, 135o 43’ 29.7”E, BVT, 26 October 2009.
Type material. Holotype. Male deposited in South Australian Museum, Adelaide, length 4.9 mm, height 2.9 mm, registration number SAM C8447.
Allotype. Female deposited in South Australian Museum, Adelaide, length 5.8 mm, height 53.9 mm, registration number SAM C8448.
Paratypes. Five males 5.0 x 3.1 mm, 5.0 x 3.0 mm, 4.2 x 2.7 mm, 4.2 x 2.4 mm, 4.0 x 2.7 mm; five females 5.7 x 3.8 mm, 5.7 x 3.6 mm, 5.3 x 3.6 mm, 5.0 x 3.2 mm, 5.0 x 3.1 mm, registration number SAM C8449
Other material. South Australia, Pygery Rocks, 7 km N of Wudinna, 32o 59’ 09.8”S, 135o 28’ 14.3”E, 20 March 2013, BVT, AM P97824; Pildappa Rocks, 13.6 km NNE of Minnipa, 32o 45’ 04.6”S, 135o 13’ 51.3”E, 20 March 2013, BVT, AM P97825; Victoria, Koorooya State Park, 15.5 km WSW of W of Inglewood, East walking track, 36o 35’ 36.2”S, 143o 42’ 48.1”E, 7 April 2014, BVT, AM P97826; Kooyoora State Park, 15.5 km WSW of Inglewood, Kirwan Rd, 36o 34’ 59.3”S, 143o 42’ 33.1”E, 8 April 2014, BVT, AM P97827; Western Australia, Yanneymooning Rocks, 40 km NE of Mukinbudin, 30o 42’ 47”S, 118o 33’ 23”E, 31 March 2003, M. Davis, WAM C59038; Elachbutting Rock, 52 km NE of Mukinbudin, 30o 35’ 22”S, 118o 36’ 41”E, 1 April, 2003, M. Davis, WAM C59039.
Diagnosis. Egg with 40 to 80 polygons with an elongated central depression, frilled edges and grainy surface.
18 trunk segments. Male with an asymmetrical rostrum subequal in size to the ocular tubercle, and clasper with palps 2– 3 x longer than the palm and with distal palpomere usually incompletely divided into two. Telson with about 10 spines somewhat unevenly spaced and sized and with 8–12 setae of variable size, but 1–1.5x diameter of cercopod.
Description. Egg ( Fig 9 View FIGURE 9 B.C.D) spherical ca.190 Μm diameter (range 186–200 Μm; n =5). About 50 polygons, each with an elongated central depression, dividing ridges frilled and with short protrusions at ridge junctions. Whole surface grainy or spaced micropores.
Male. Head ( Fig 11 View FIGURE 11 C) with prominent ocular tubercle with round compound eye occupying most (ca 70%) of it. Rostrum extended asymmetrically at about an angle of 110o to form a protrusion about the same size as the ocular tubercle. Ocellus about half the size of the ocular tubercle and located in the middle base of the rostrum. Dorsal organ posterior to the eye by about its height, pedunculate and with a flattened apex at a 45o angle to the peduncle and only about two-thirds the ocular tubercle height.
First antennae a little longer than peduncle of second antennae and with about 7 lobes each with numerous tiny sensory setae.
Second antennae with spinose peduncle and each flagella with about 8 antennomeres, dorsally with 1–4 spines and ventrally with 2–5 long setae. Dorsal spines less numerous in basal and two most distal antennomeres, and most antennomeres with 3–4 ventral setae, with lesser numbers on the basal antennomere and more on the 2–3 distalmost antennomeres.
Carapace ( Fig 11 View FIGURE 11 A) elongated oval, pellucid with little indication of growth lines. Dorsal edge at about one quarter its length with a slight depression where body is attached to the carapace and adjacent to the adductor muscle scar lying at about 45o to the horizontal body axis.
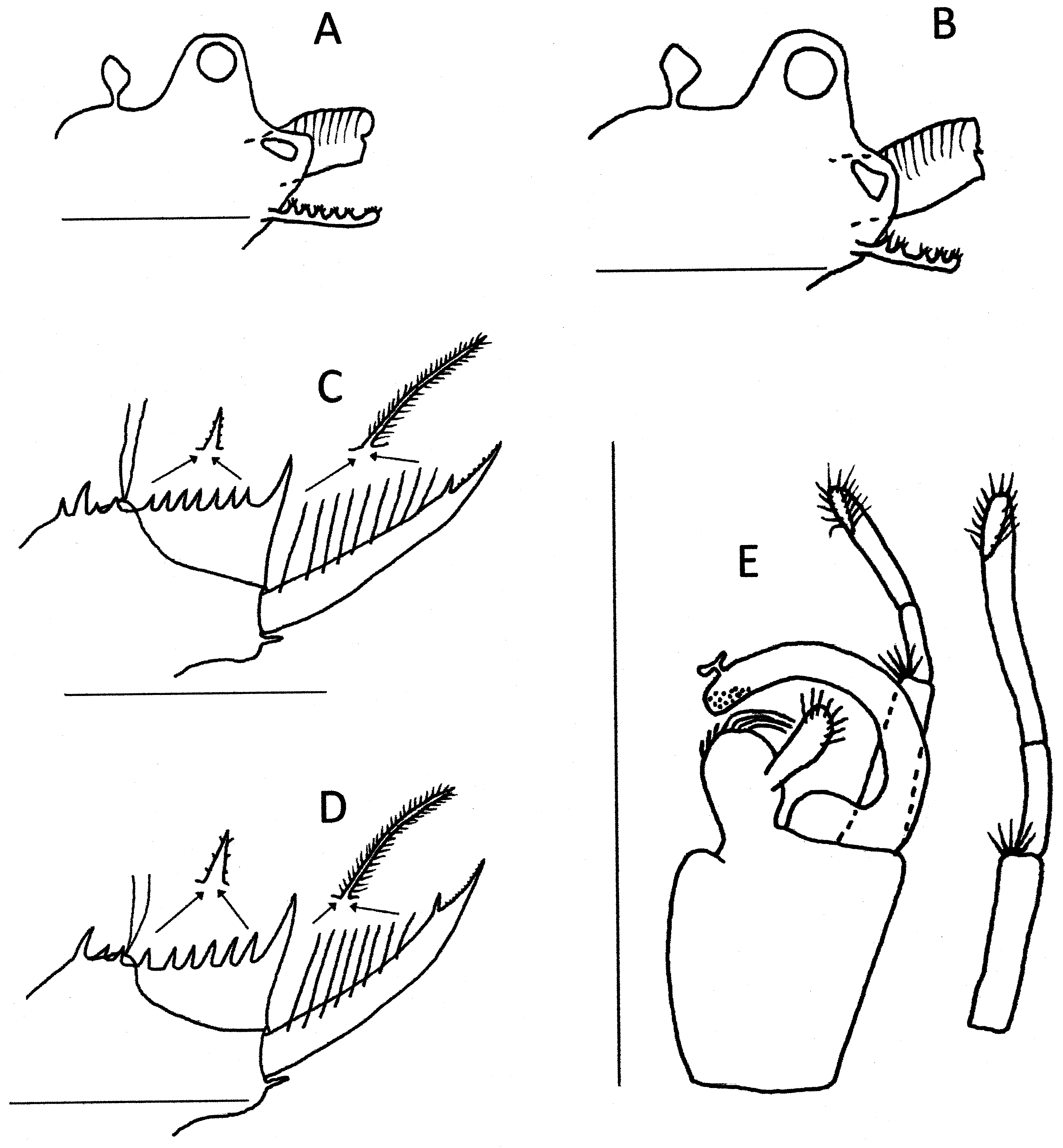
Thoracopods. Eighteen pairs, the first two modified as claspers ( Fig 11 View FIGURE 11 G). The first clasper with palm trapezoidal but with a small rounded projection medodistally, apical club rounded with thick denticles distomedially and spines apicolaterally, movable finger of normal curved structure. First clasper with palp 2x palm length, second clasper with palp about 2.5x palm length. Five short spines located medially at junction of the palpomeres, and distal palpomere incomplete subdivided, so that they appear 3 segmented.
Other thoracopods of typical structure for Eulimnadia , decreasing is size and complexity posteriorly. Dorsal surface of trunk with 1–3 short spines posteriorly on each of the posterior 8 trunk segments.
Telson ( Fig 11 View FIGURE 11 E) with about 11 pairs of dorsal spines, well spaced, the first the largest. Most spines with denticles. Caudal filaments originating from a mound a little higher than the dorsal floor of the telson and between the 3rd and 4th spine. Dorsal floor of telson with a sharp declivity immediately posterior to the mound, followed by a slow decline to the base of the cercopod. Cercopods a little longer than dorsum of telson, the basal three-quarters hardly thinning to a small spine with denticles, then a rapid thinning to an acute apex. About 12 setae, the first few shorter, subequal to cercopod basal diameter, and almost naked while most longer, about 1.5x cercopod diameter and with both segments well feathered. A cirrus of many tiny denticles dorsolaterally on apical quarter. Prominent spiniform projection beneath the cercopods at the ventroposterior corner of the telson.
Female. Head with ocular tubercle prominent, with compound eye occupying most (ca 70%) of it. Rostrum a smooth bulge at an angle of about 150o to the frons and with the centrobasal part occupied by an ocellus, and subequal in size to the ocular tubercle. Dorsal organ posterior to eye as in male.
First antennae subequal in length to peduncle of the second antennae, with four small lobes each with many short sensory hairs.
Second antennae as in male.
Carapace as in male, though dorsally more vaulted.
Thoracopods. Eighteen of typical Eulimnadia structure.
Trunk dorsum with 3–9 setae terminally, these setae few, short and stout on last few segments, numerous and longer on segments 8–15 and hardly any setae on anterior trunk segments 1–7.
Telson ( Fig 11 View FIGURE 11 F) as in male, though fewer cercopod setae, though same arrangement with a few shorter ones basally and about 5 longer ones distally.
Variability. Populations across southern Australia vary in some details. Those from Kooyoora State Park in central Victoria ( Fig 12 View FIGURE 12 ) have similar male and female heads, though the latter is not as protrusive, being about half the size of the ocular tubercle. There are about 11 telsonic denticles in each sex, with a few specimens having an extra spine anterior to the larger first one. The cercopod setae number about 9 (range 8–10) and are less variable in size than in the type specimens, but 1– 2 x cercopod diameter in length. The clasper differs slightly in having the distal palpomere of large palp usually completely subdivided. Eggs ( Fig 9 View FIGURE 9 B) have about 40 polygons of similar structure as in the type material.
The Western Australian (Yanneymooning & Elachbutting) material ( Fig 13 View FIGURE 13 ) is also slightly different from the type material. Heads are similar, though again the female is less protruded. The second antenna has only 7 antennomeres. The large palp of the clasper has a spine midlaterally on the basal palpomere and the distal palpomere is undivided. There are about 10 telsonic spines, even more spaced particularly anteriorly, than the Peela material and almost always with a small supernumerary anterior to the first large spine. As in the Peela specimens, cercopod setae number about 12 and are variable in size, with about 2 smaller ones basally. Lengths vary from about 1x to 1.5x cercopod diameters. Eggs ( Fig 9 View FIGURE 9 D) have about 70–80 polygons, but otherwise they are of similar structure to those from Victoria.
Comments. Eulimnadia gnammaphila sp. nov. has a wide distribution (see below) but there is sufficient similarly between the populations to accommodate all in one morphological species. It is possible future studies using molecular techniques will be able to provide finer differentiation. In particular the Western Australian material could prove to be specific with its egg having far more polygons and the clasper palp having a spine on the basal palpomere and the distal palpomere being whole. In addition, the second antenna has only seven antennomeres and this does not appear to be due to accidental breakage.
As in many species of Eulimnadia , hermaphrodites (which look like females) outnumber males in most collections, often by 10:1 or more. The Pygery collections lack males while those from Kooyoora have about 50% males, but these are biased as males were targeted in the field. In the Elachbutting sample the ratio is 20 females: 4 males, and in the Yanneymooning sample 21:10. Reproduction is probably androdioceous (Weeks et al., 2008).
Distribution and ecology. Eulimnadia gnammaphila sp. nov., as presently defined, occurs in granitic gnammas across southern Australia, from central Victoria to the wheatbelt of southern Western Australia. It was earlier misidentified (see Timms, 2015 and this paper) variously as E. feriensis and E. dahli in the wheatbelt (Weeks et al., 2006). Generally it occurs for only a few weeks in the pools after they fill after storm rains in summer or after the first rains in autumn. Although widespread, E. gnammaphila sp. nov., is uncommon, as apparently only a few pools on some rock outcrops have an egg bank, and it is absent during much of the filling cycle in winterspring ( Timms, 2014).


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |