
Persicolaelaps, Kazemi, Shahrooz & Beaulieu, Frédéric, 2016
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4200.4.2 |
|
publication LSID |
lsid:zoobank.org:pub:B787F2B5-60FB-4E29-89D9-16C544200E89 |
|
DOI |
https://doi.org/10.5281/zenodo.6085649 |
|
persistent identifier |
https://treatment.plazi.org/id/BF7057AB-8686-4230-83CB-5E6BEFBEE18D |
|
taxon LSID |
lsid:zoobank.org:act:BF7057AB-8686-4230-83CB-5E6BEFBEE18D |
|
treatment provided by |
Plazi |
|
scientific name |
Persicolaelaps |
| status |
gen. nov. |
Persicolaelaps new genus
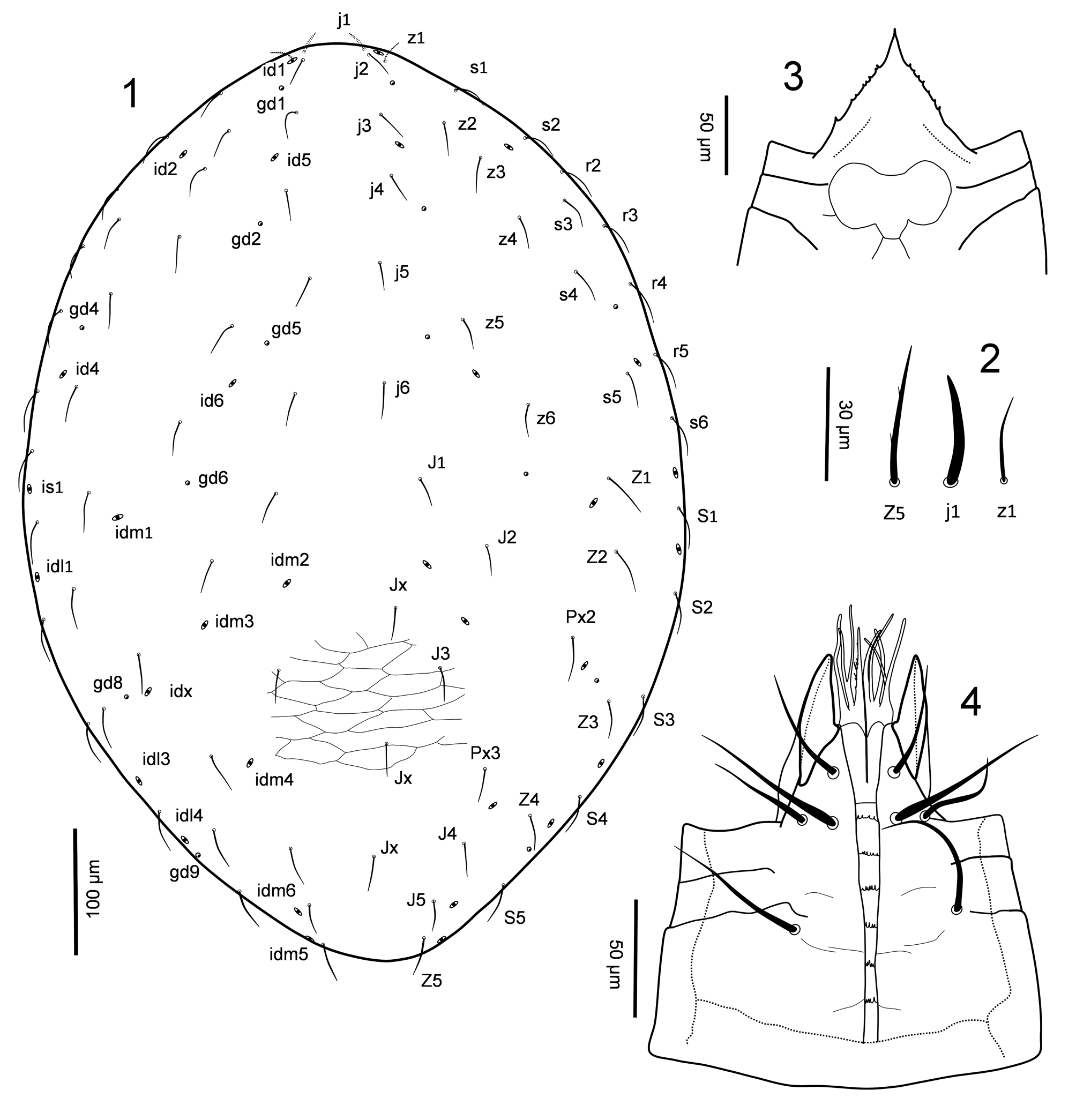
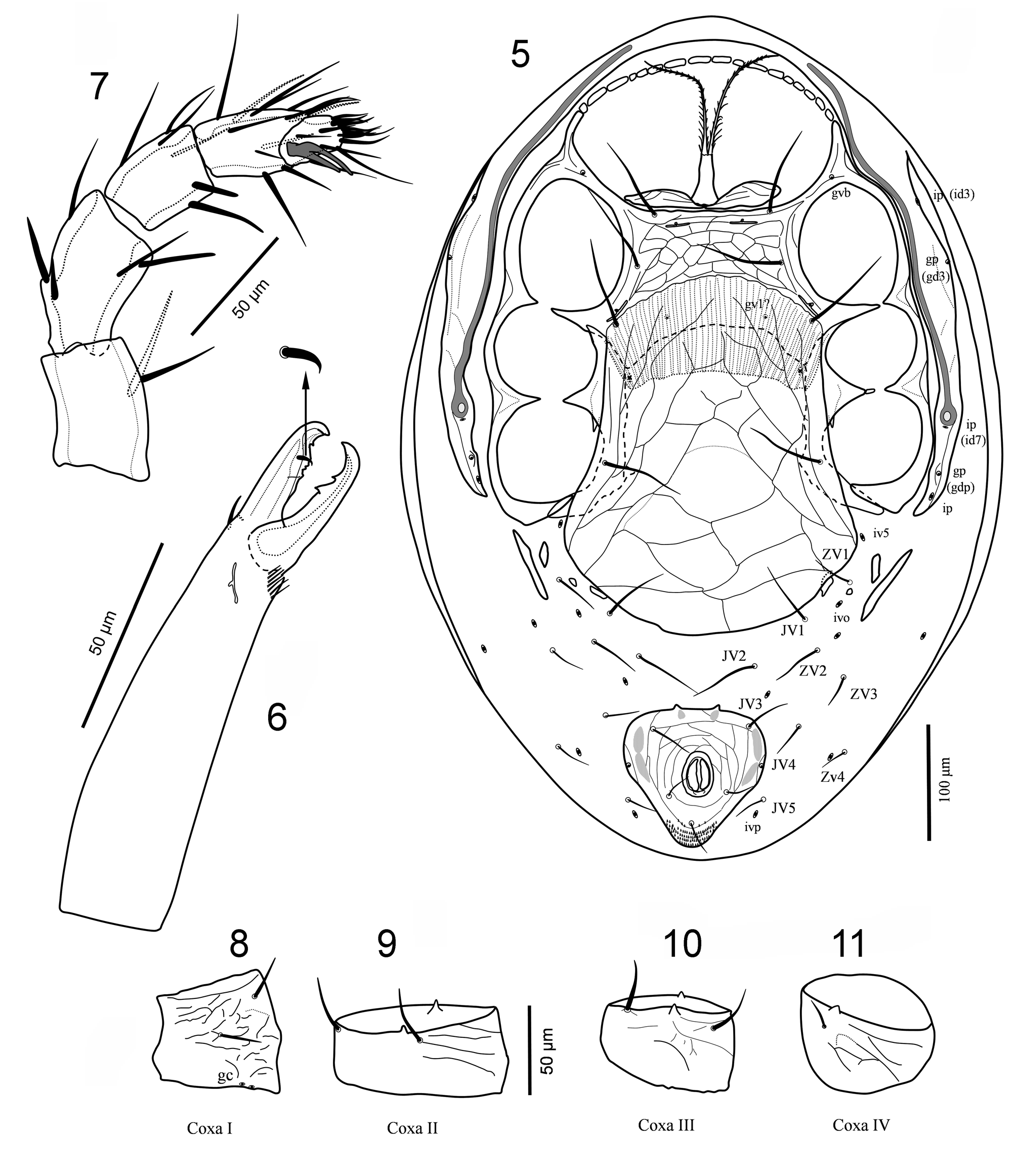
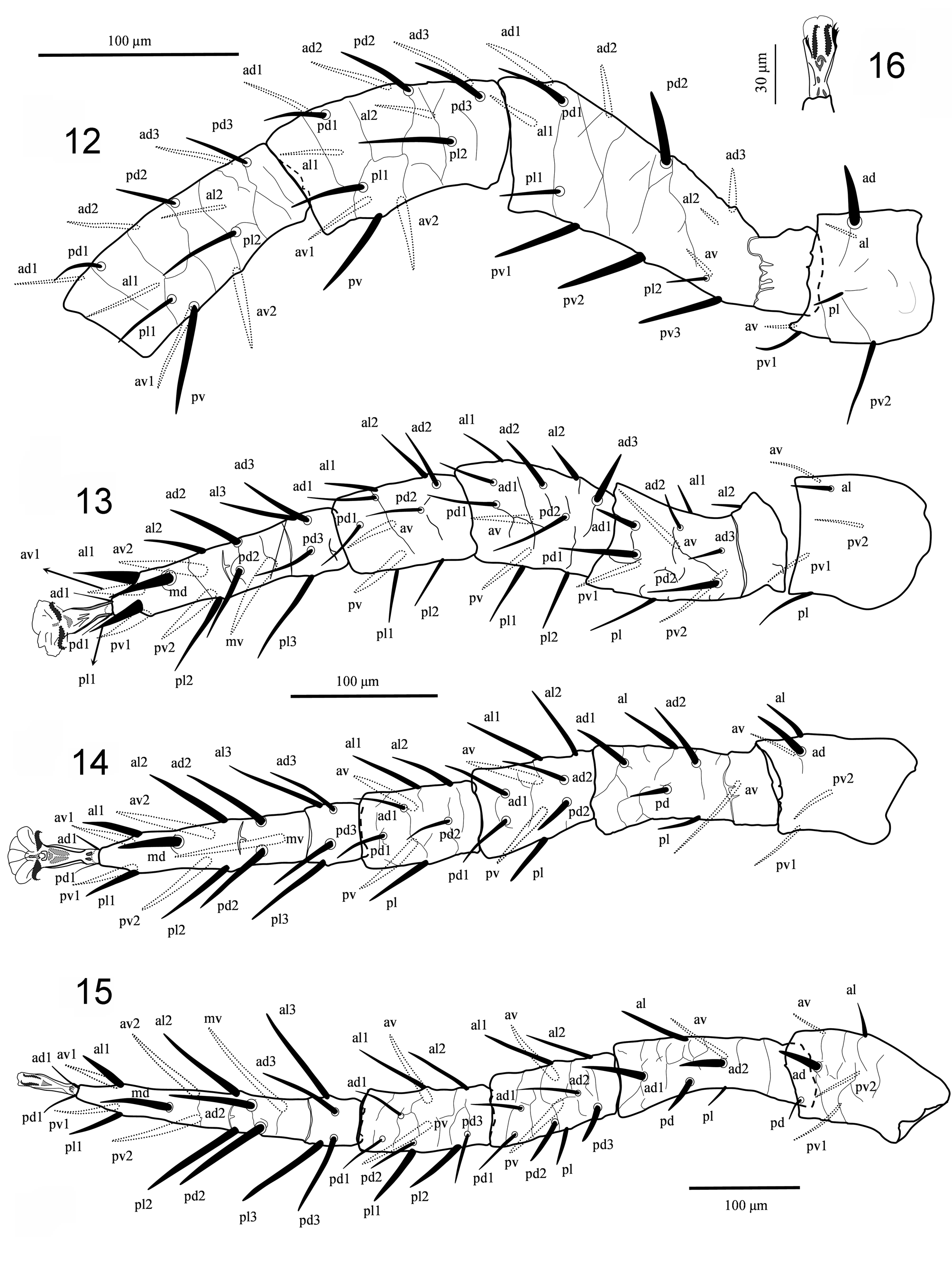
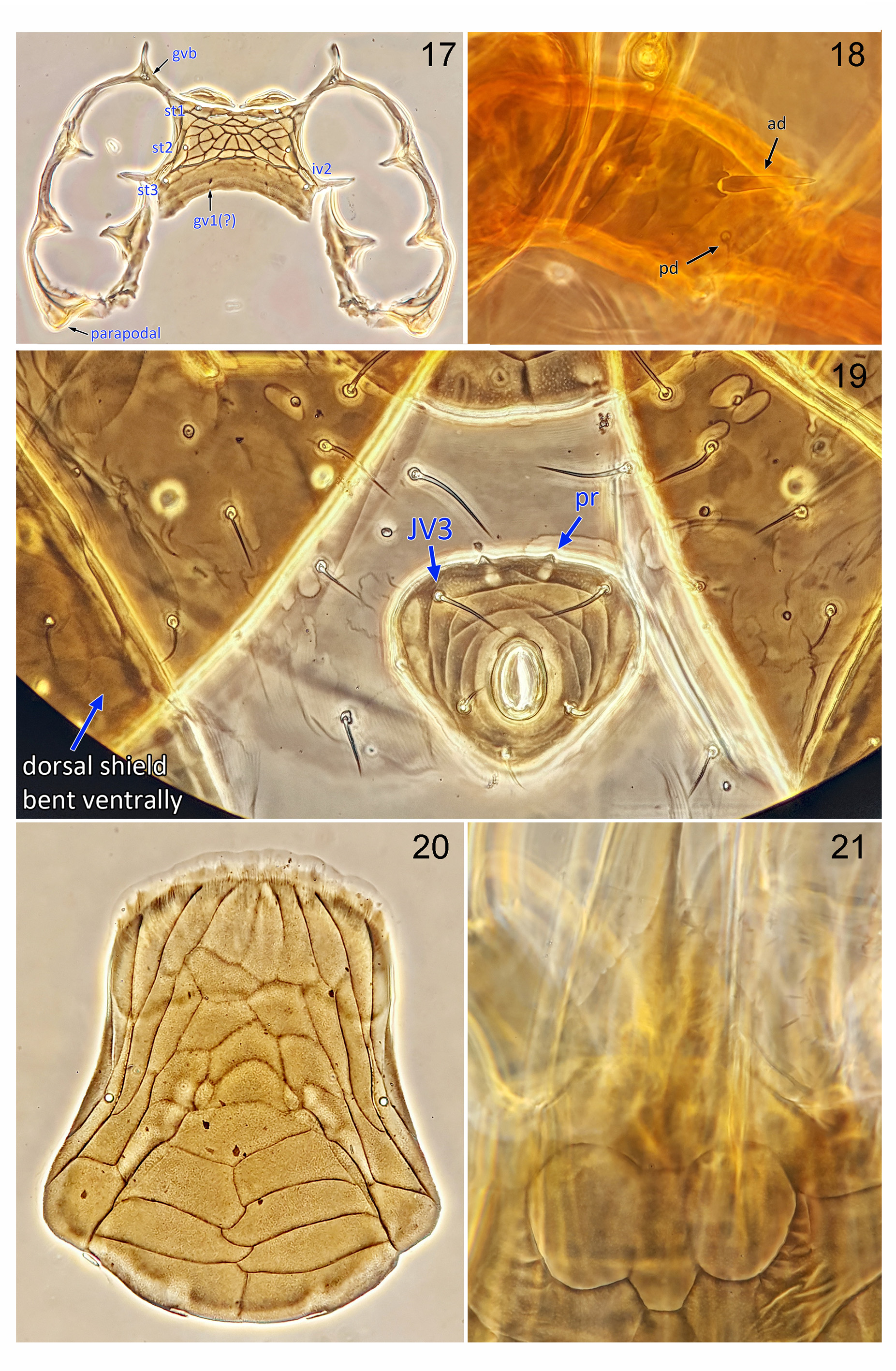
( Figs 1–21 View FIGURES 1 – 4 View FIGURES 5 – 11 View FIGURES 12 – 16 View FIGURES 17 – 21 )
Type species: Persicolaelaps hallidayi sp. nov., by monotypy.
Diagnosis (adult female). The genus can be readily distinguished from other laelapids by the following characters, unique or rarely observed in the family: (1) an epigynal shield widened and expanded anteriorly, overlapping much of posterior half of sternal shield, axe-shaped posteriorly; well removed from anal shield, bearing st5. Anteroventral region of idiosoma well sclerotised, showing (2) fusion of sternal shield with endopodal and exopodal plates between acetabula I and II, and (3) a strip of small subrectangular sclerites extending from anterior extension of exopodals and flanking dorsally coxae I and gnathosomal base. (4) An anal shield normally shaped (inversely subtriangular), capturing one pair of ventral setae (JV3). (5) Metasternal setae st4 absent. Legs with (6) trochanter IV bearing six (1 1/1 1/2 0) setae, including pd, and (7) genu IV bearing 10 setae (2 2/1 3/1 1), including pv (this seta is present in a few other laelapid genera); leg setation otherwise normal for Laelapidae (sensu Evans 1963b). (8) Gnathotectum triangular, acuminate, finely denticulate laterally. (9) Internal malae composed of several thick, tentacle-like projections. The genus is further defined by this combination of characters variously shared with other Laelapidae : dorsal shield covering all dorsal idiosoma, oval, somewhat dome-like, bearing 39 pairs of setae, including px2–3, and a few unpaired setae Jx; dorsal setae short, mostly smooth and slender; r6 and R series absent; shield with 23 pairs of pore-like structures, including seven pairs of gland pores (gd1–2, gd4–6, gd8–9). A pair of well-sclerotised presternal platelets. Sternal shield well sclerotised, wider than long. Peritremes long, narrow. Deutosternal groove moderately narrow, with six rows of denticles, each with 2–6 denticles. Chelicera moderately stout, chelate-dentate; pilus dentilis normal, short, setiform. Palp tarsal claw two-tined.
Description. Dorsal idiosoma ( Fig. 1 View FIGURES 1 – 4 ). Dorsal shield covering all dorsal idiosoma, oval-shaped, dome-like, slightly extending to venter laterally, bearing 39 pairs of short, slender setae ( Fig. 2 View FIGURES 1 – 4 ); setae Jx may be present; r6 and R series absent. Dorsal shield with a complement of 23 pairs of pore-like structures, including seven pairs of gland pores (gd1–2, gd4–6, gd8–9) and 16 pairs of poroids.
Ventral idiosoma ( Fig. 5 View FIGURES 5 – 11 ). Tritosternum with a pair of free pilose laciniae and columnar base. A pair of welldeveloped sclerotised presternal platelets. Sternal shield wider than long ( Figs 5 View FIGURES 5 – 11 , 17 View FIGURES 17 – 21 ), well sclerotised, fused to endopodal elements between coxae I–II into arms that join exopodal strips; a series of small sclerites extending from anterior extension of podal elements and dorsally flanking gnathosomal base and coxae I ( Fig. 5 View FIGURES 5 – 11 ). Metasternal setae st4 absent; iv3 on soft cuticle. Epigynal shield ( Fig. 20 View FIGURES 17 – 21 ) expansive, broadened posteriorly, angled posterolaterally behind coxae IV, rounded posteromedially, well separated from anal shield, lineate-reticulate, with a wide Λ-shape lineation, bearing pair of epigynal setae (st5). Anal shield inversely subtriangular ( Fig. 19 View FIGURES 17 – 21 ), capturing one pair of ventral setae (JV3). Peritrematal shield well developed, widely fused anteriorly to dorsal shield, free posteriorly, bearing five discernible pairs of pore-like structures, including two gland pores and three poroids. Endopodal platelets between coxae III–IV well developed, free from sternal shield, although may overlap it. Opisthogastric region not hypertrichous.
Gnathosoma ( Figs 3–4 View FIGURES 1 – 4 , 6–7 View FIGURES 5 – 11 , 21 View FIGURES 17 – 21 ). Gnathotectum a subtriangular, acuminate projection, denticulate laterally ( Figs 3 View FIGURES 1 – 4 , 21 View FIGURES 17 – 21 ). Corniculi horn-like; internal malae longer than corniculi, composed of a pair of fringed median projections, and lateral tentacle-like projections; hypostomal and capitular setae smooth; deutosternal groove with six rows of denticles, and a smooth ridge anteriorly ( Fig. 4 View FIGURES 1 – 4 ). Chelicera moderately stout, chelate-dentate, with relatively thick, setiform dorsal seta, fixed digit with few small teeth, pilus dentilis short and setiform, movable digit bidentate ( Fig. 6 View FIGURES 5 – 11 ). Palp chaetotaxy normal for Gamasina (sensu Evans, 1963a) with 2-5-6-14 -15 setae on trochanter-femur-genu-tibia-tarsus; tarsal claw two-tined ( Fig. 7 View FIGURES 5 – 11 ).
Legs ( Figs 8–16 View FIGURES 5 – 11 View FIGURES 12 – 16 , 18 View FIGURES 17 – 21 ). Legs moderately long, leg I approximately as long as idiosoma, leg IV longer than idiosoma; all legs with well-developed paired subequal claws and rounded pulvilli borne on ambulacral stalk. Setation of legs I-II-III-IV: coxae 2, 2, 2, 1; trochanters 6, 5, 5, 6, trochanter IV with a pd seta (1 1/1 1/2 0) ( Fig. 18 View FIGURES 17 – 21 ); femora 13 (2 3/1 2/3 2), 11 (2 3/1 2/2 1), 6 (1 2/1 1/0 1), 6 (1 2/1 1/0 1); genua 13 (2 3/2 3/1 2), 11 (2 3/1 2/1 2), 9 (2 2/1 2/1 1), 10 (2 2/1 3/1 1) ( Fig. 20 View FIGURES 17 – 21 ); tibiae 13 (2 3/2 3/1 2), 10 (2 2/1 2/1 2), 8 (2 1/1 2/1 1), 10 (2 1/1 3/1 2); tarsi II-IV 18 (3 3/2 3/2 3 + mv, md). Many setae on tarsi II–IV slightly thickened or spine-like, especially ventral setae, dorsal and ventral setae of femur I, and ventral setae of genua and tibiae ( Figs 12–15 View FIGURES 12 – 16 ).
Remarks. In addition to the attributes mentioned in the introduction, the well-developed internal malae of Persicolaelaps are indicative of Laelapidae in general, and their thick, elongate filaments show a resemblance (or possible relation) to some laelapids species such as Cosmolaelaps cuneifer (Michael) ( Evans & Till, 1966) , Cosmolaelaps vacua species group (Michael), Laelaspis spp., Pneumolaelaps spp. (FB & SK pers. obs.), Androlaelaps walkerae Till, 1963 , Gaeloaelaps saboori Joharchi & Babaeian, 2014 and ‘ Hypoaspis’ cavitatis Karg, 1982, (which, at least superficially, resemble some Laelaspisella species). The internal malae of Mesolaelaps accessoria Tenorio & Radovsky, 1974 also have thick projections, but they appear shorter and originate from the lateral margin of the main, median processes. Some Laelaspisella and the apparently related Pogonolaelaps species have elongate fringed internal malae, but composed of hair-like and thinner filaments, based on illustrations and personal examination of some species ( Marais & Loots, 1969; Nemati & Gowiazdowicz, 2016; SK pers. obs.).
Persicolaelaps actually shares most of the character states of the putative early-derivative lineage of dermanyssoids, best exemplified by some members of Gaeolaelaps or the ‘ Hypoaspis complex’ ( Laelapidae View in CoL ), from which other groups of facultative and obligate haematophagous groups of laelapids and other dermanyssoids theoretically evolved ( Evans & Till, 1966; Radovsky, 1969, 1994). These basal laelapids are part of the subfamily Hypoaspidinae, which includes a large portion of the free-living (putatively predatory) laelapids and many insect symbionts. Like many hypoaspidines, Persicolaelaps also has a strong leg chaetome, with many setae thickened or spine-like. Persicolaelaps can suitably be placed in this subfamily, until laelapid classification improves to better reflect phylogenetic relationships. The departure of Persicolaelaps from the basic laelapid or dermanyssoid type (see Evans & Till, 1966: 111) corresponds to the main diagnostic features of the genus (features #1–9 listed in genus diagnosis), as well as the addition of dorsal unpaired setae J, the loss of setae r6, R and UR, and a dorsal shield that covers the idiosoma entirely. However, each of these features also occur in at least a few other members of laelapids, with the exception of the anal shield bearing setae JV3, which may well be unique in Laelapidae View in CoL , and trochanter IV with six setae, which appears as a unique attribute within Mesostigmata View in CoL ( Evans, 1963a, 1972). The fragmented band of sclerites extending anteriorly from fused endo-exo-sternal elements is also exceptional, and may be rare in Mesostigmata View in CoL . Note that other laelapids, e.g. Pseudoparasitus jilensis Ma, 2004 and Urozercon robustisetae Rosario & Hunter, 1988 , and other dermanyssines (some Veigaiidae, Evans & Till, 1979 View in CoL ) also have a normally-sized anal shield bearing five setae, including a pair of opisthogastric setae in addition to the circumanals. Some Haemogamasus species have several additional setae on a normally-sized anal shield, but this is due to strong opisthogastric hypertrichy ( Evans & Till, 1966).
A number of genera that are generally classified as hypoaspidines (e.g. Vitzthum, 1940 –1943: 762; Karg , 1965) share several of the diagnostic features of Persicolaelaps , which in some cases, might reflect a phylogenetic relationship, or alternatively, morphological convergence, or even plesiomorphies. In particular, species of Ololaelaps share with Persicolaelaps a strongly developed, well-sclerotised (thick, as indicated by the brownish tint), rounded dorsal shield (more elongate in some Ololaelaps, Jordaan & Loots, 1987 , SK pers. obs.), bearing attenuate (slender) dorsal setae; an epigynal shield broadly expanded anteriorly; a pair of sclerotised presternal platelets; endopodals, exopodals and sternal shield more or less coalesced anteriorly; well-developed endopodals mesad coxae III–IV; relatively broad peritrematal shield; parapodal plate thicker than usual; a gnathotectum somewhat triangular (less acuminate in Ololaelaps ), finely denticulate (more sparingly in Ololaelaps ); and a deutosternal groove with few denticles per row ( Evans & Till , 1966; Jordaan & Loots , 1987; FB & SK, pers. obs. on Ololaelaps spp. from North America , Iran and Australia). Furthermore , at least some Ololaelaps have a band of sclerotised cuticle extending anteriorly from the podal plates between coxae I and II, and flanking coxae I laterally ( Jordaan & Loots, 1987; we have seen such a cuticular band, although weakly sclerotised, in some specimens of at least two species examined by us, FB & SK). It is possible that this band extends and dorsally surrounds the gnathosomal base (but observation is difficult in this dark, crowded region), just as the series of (more conspicuous) sclerites do in Persicolaelaps . Ololaelaps is otherwise readily distinguished from Persicolaelaps by its epigynal shield hyperdeveloped posteriorly and fused to the anal shield, the presence of setae st 4 in adult female, more slender leg setae, as well as by not having several other features characterising Persicolaelaps (#2, 4, 6, 7, 9 in genus diagnosis above).
Although also exhibiting many differences from Persicolaelaps , species of Oloopticus , Laelaspisella, Pogonolaelaps and Reticulolaelaps share with it a salient attribute: the absence of metasternal setae ( Marais & Loots, 1969; Karg, 2003; Nemati et al., 2013; Nemati & Gwiazdowicz, 2016; Joharchi et al., 2016). Except for Oloopticus , which has a well-developed sternal shield (and apparently bearing deep pits at the region usually occupied by st4), the absence of st 4 in these genera and Persicolaelaps may be a result of a marked anterior expansion of the epigynal shield, leaving too little space, and little functional role (being so close to another pair of setae, st3) for them to develop. An intermediate stage may be exhibited by Ololaelaps , for which seta st4 is inserted in the posterolateral margins of an otherwise normal sternal shield, near setae st3. Setae st4 are also lost in some species of the insect-associated laelapid genera Myrmozercon , Urozercon and Dinogamasus ( Lundqvist, 1998; Klimov & OConnor, 2004; Ghafarian et al., 2013). The sternal shield is also broader than long in Ololaelaps , Laelaspisella and Pogonolaelaps, and concave posteriorly (also in Reticulolaelaps ), seemingly also to accommodate the anterior expansion of the epigynal shield, a correlation seen in some members of many other genera of laelapids or relatives, especially associated with insects (e.g. Pneumolaelaps , Holostaspis , Laelaspis , Bisternalis , insect-associated Gaeolaelaps , Gymnolaelaps myrmophila (Michael) , Melittiphisoides , Melittiphis alvearius (Berlese) ; Evans & Till, 1966; Baker et al., 1983; Delfinado-Baker et al., 1984; Kazemi et al., 2014) or vertebrates ( Androlaelaps , Haemogamasus ; Till, 1963) and more strikingly in dermanyssids, macronyssids and hirstionyssids ( Evans & Till, 1966; Domrow, 1988), for which this may represent an adaptation for an increased capacity for engorgement in blood by having the opisthogastric region mostly covered by soft cuticle (Radovsky, 1969).
Laelaspisella, Pogonolaelaps and Reticulolaelaps share an additional feature with Persicolaelaps : the presence of two ventral setae on genu IV. This character also occurs in several disparate laelapid genera (e.g. Pneumolaelaps , Melittiphis , and some Laelaps and Myrmozercon species, Shaw & Seeman, 2009), as well as several genera of macronyssids ( Evans & Till, 1965), and a few species of ‘ Hypoaspis’ s. lat. that are difficult to classify (e.g. H. giffordi Evans & Till , H. lubrica Voigts & Oudemans , H. atomarius Berlese ; Evans & Till, 1966; van Aswegen & Loots, 1970). This indicates the plasticity in the development of seta pv of genu IV. Indeed, genu IV bears a pv seta in members of various other gamasine families, while being absent in many others ( Evans, 1963a).
Etymology. The generic name Persicolaelaps refers to the ancient name of Iran, Persia, the country of origin of the genus (feminine).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Persicolaelaps
| Kazemi, Shahrooz & Beaulieu, Frédéric 2016 |
Pseudoparasitus jilensis
| Ma 2004 |
Urozercon robustisetae
| Rosario & Hunter 1988 |
Ololaelaps
| Jordaan & Loots 1987 |
Veigaiidae
| Evans & Till 1979 |