
Epimetopus Lacordaire, 1854
|
publication ID |
https://doi.org/ 10.37520/aemnp.2021.001 |
|
publication LSID |
lsid:zoobank.org:pub:9CB2C16A-E2B7-4C17-A310-D538AA061911 |
|
persistent identifier |
https://treatment.plazi.org/id/038BAE5E-FFC7-A238-F95D-FF66FE6EFB57 |
|
treatment provided by |
Carolina |
|
scientific name |
Epimetopus Lacordaire, 1854 |
| status |
|
Epimetopus Lacordaire, 1854 View in CoL
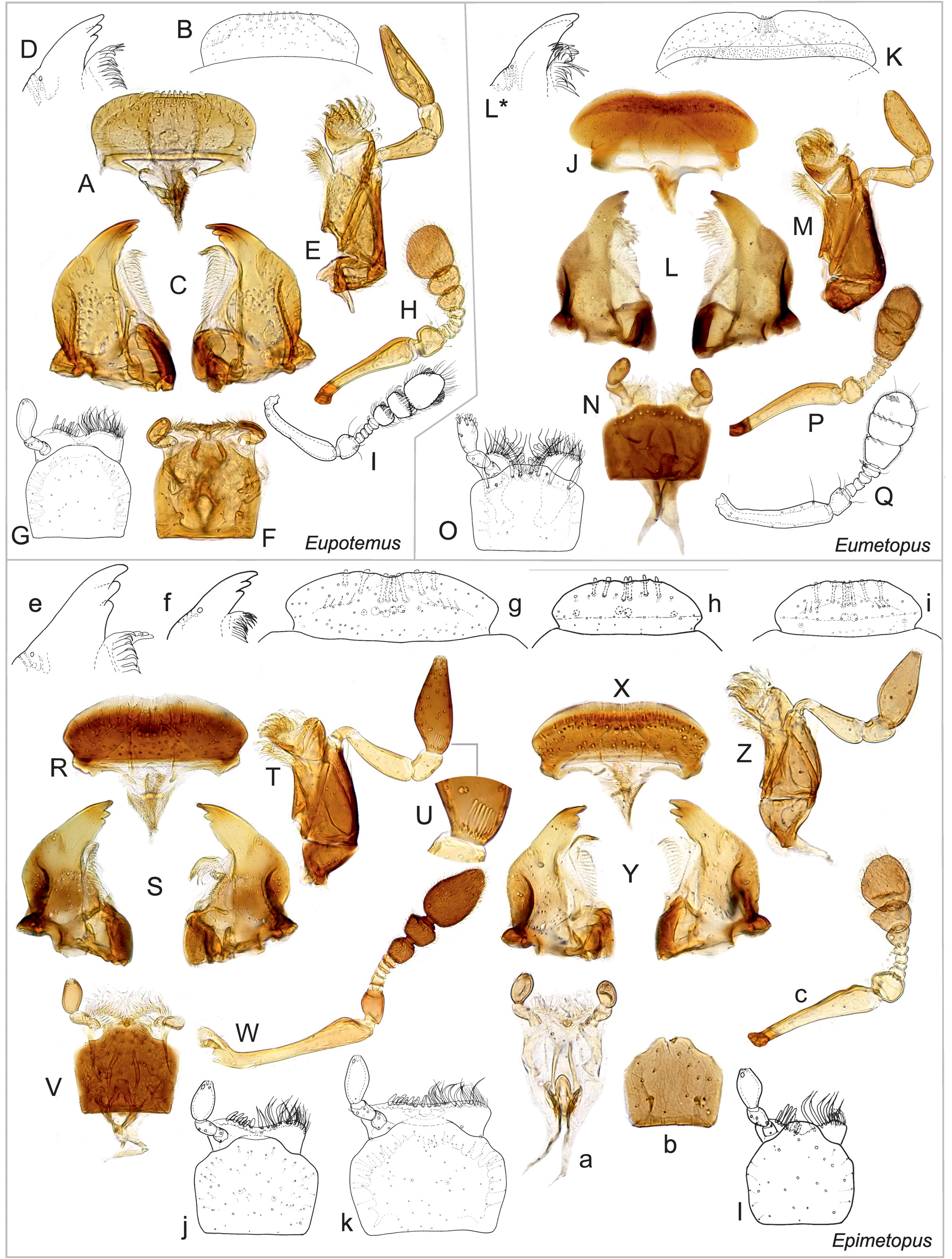
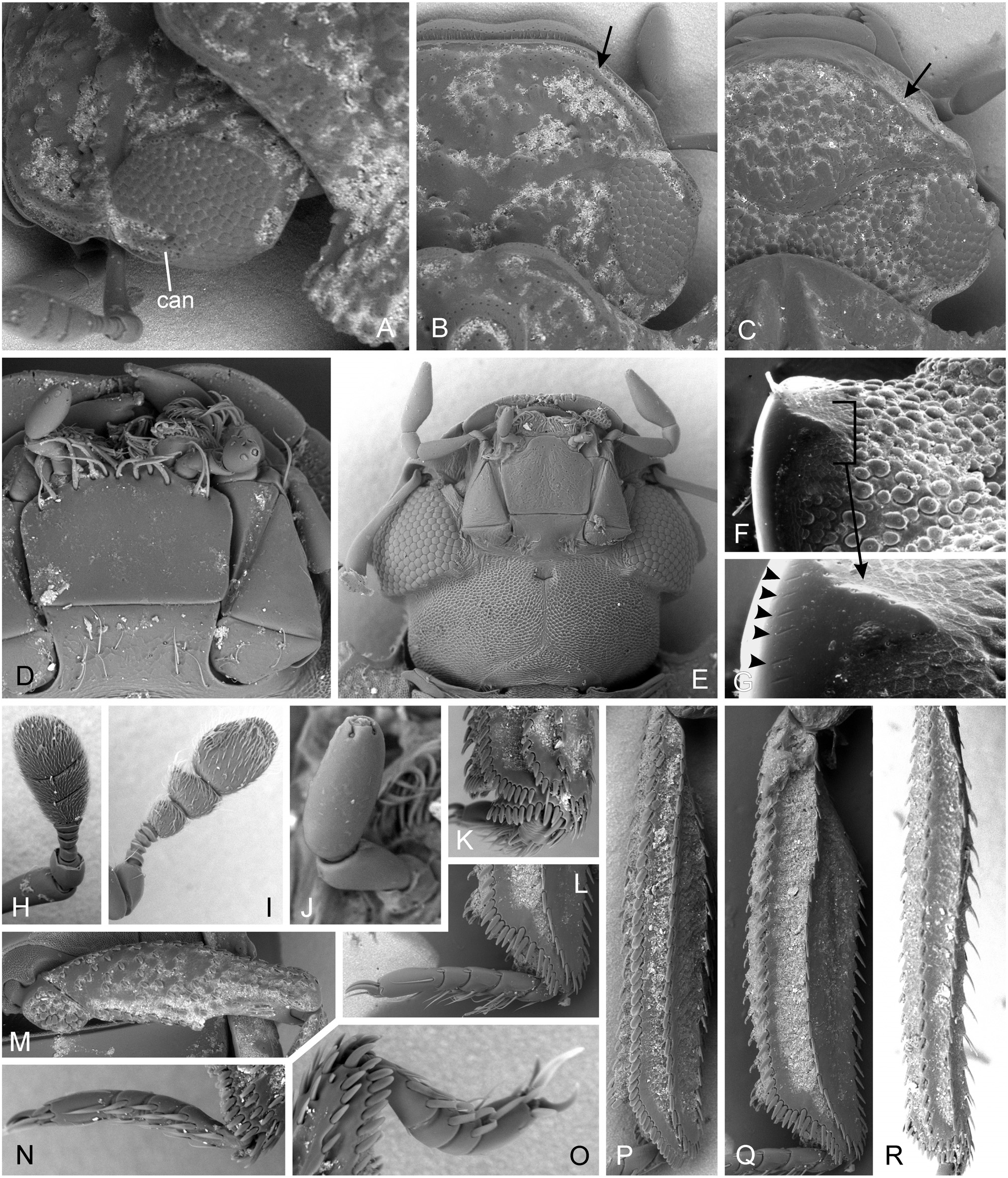
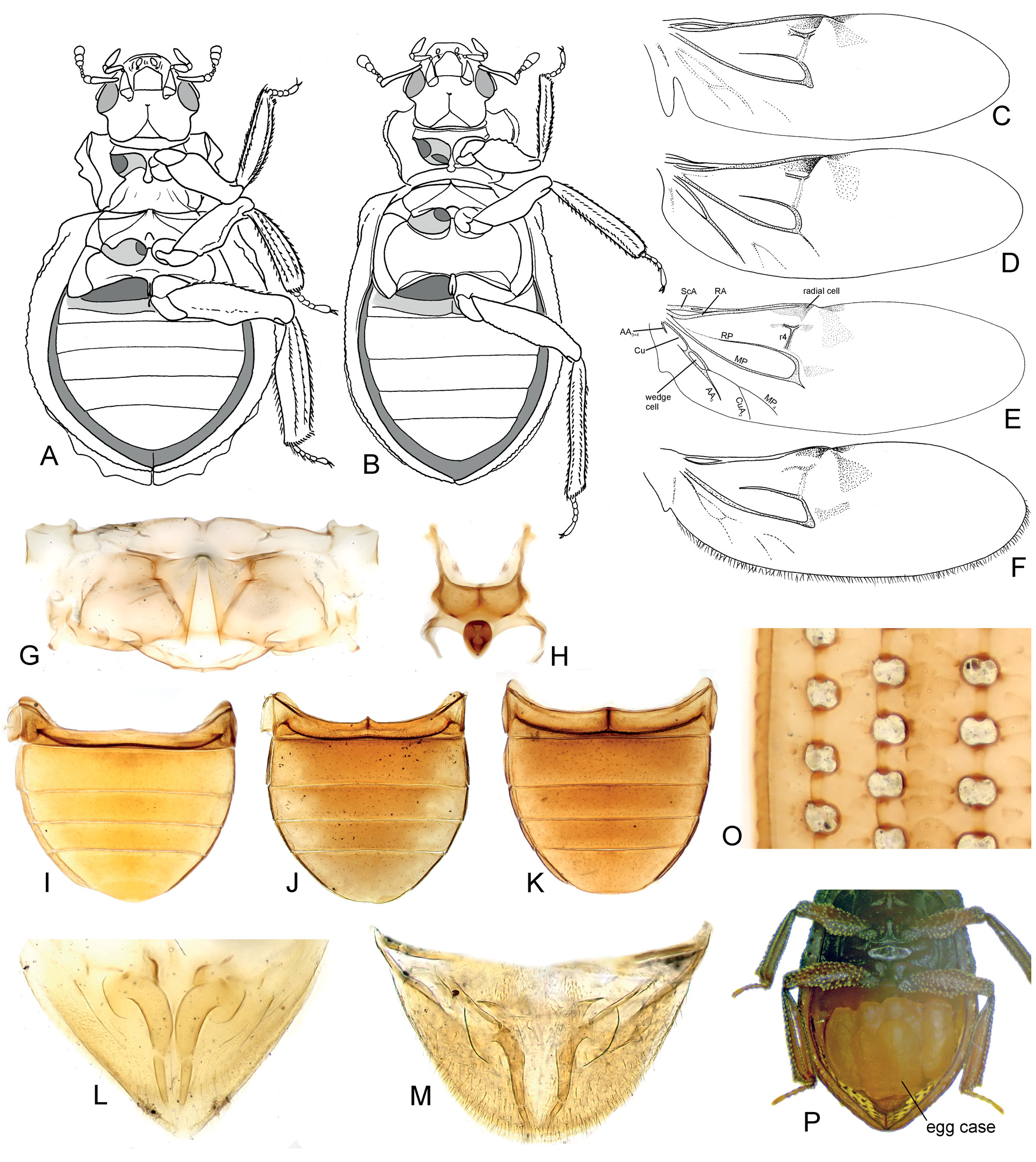
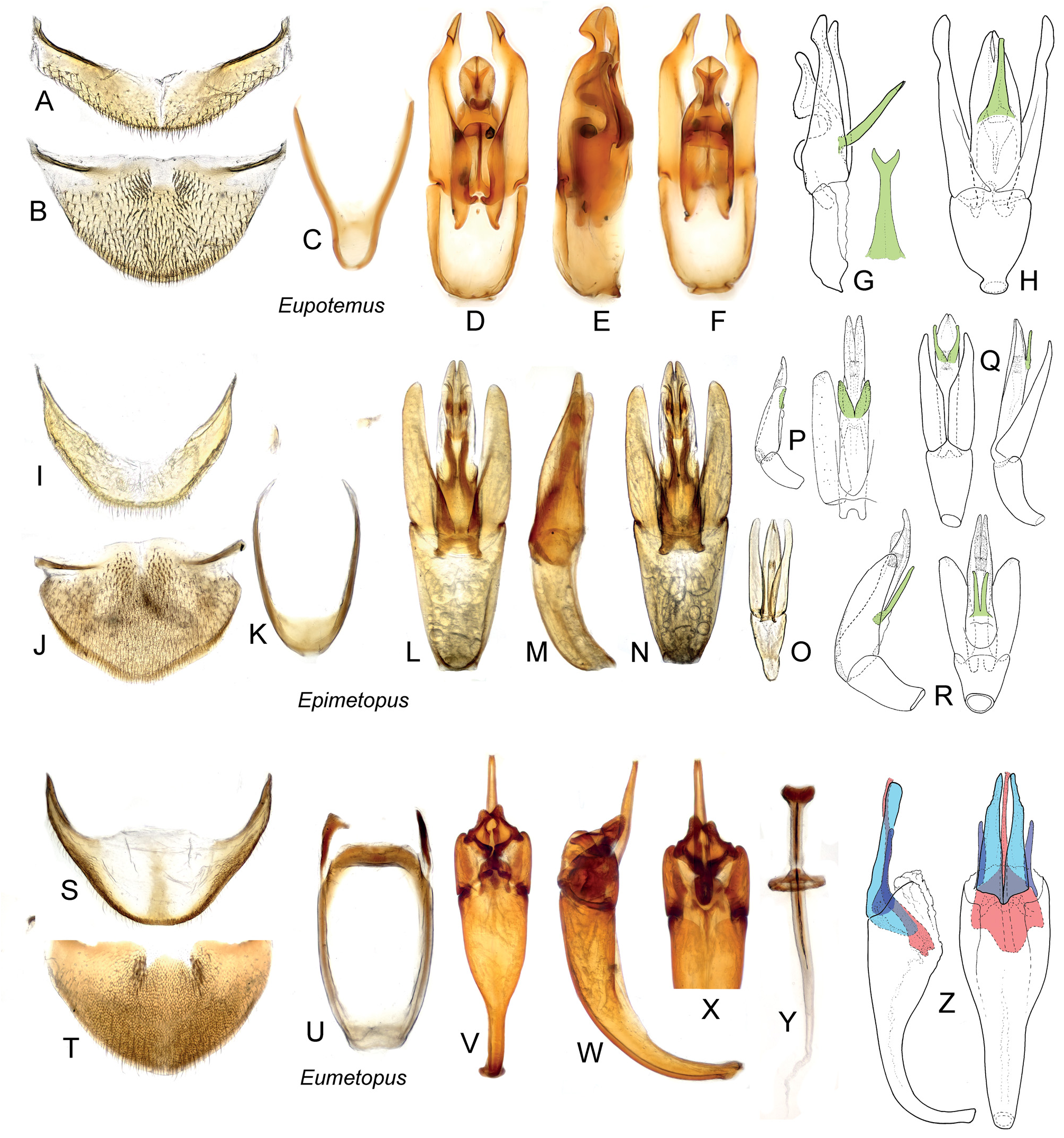
( Figs 2R–Z, a–l View Fig ; 3E–I, K, P View Fig ; 4E–G, I–J, N–P, R View Fig ;
5B, E–F, G–H, K, O–P View Fig ; 6I–R View Fig ; 16 View Fig )
Ceratoderus Mulsant, 1851: 1 View in CoL . Type species: Ceratoderus graniger Mulsant, 1851 View in CoL .
Epimetopus Lacordaire, 1854: 467 View in CoL . New replacement name for Ceratoderus Mulsant, 1851 View in CoL due to the homonymy with Ceratoderus Westwood, 1841 View in CoL .
Sepidulum Leconte, 1874:47 View in CoL . Type species: Sepidulum costatum Leconte, 1874 View in CoL ; synonymized by Hඈ*©ඇ (1876: 251).
Diagnosis. Small to moderately large species (body length 1.2–3.7 mm); body reddish to black, without metallic sheen ( Figs 16A–F View Fig ); eyes completely divided into dorsal and ventral portion ( E. trogoides group; Fංκගඹൾκ et al. 2011: fig. 12) or not (remaining groups; Fංκගඹൾκ et al. 2011: fig. 11); anterior portion of clypeus not divided from posterior parts; labrum not strongly narrowed posteriorly ( Fig. 2R, X, g–i View Fig ); mandibular apex tridentate ( Figs 2S, Y, e–f View Fig ); apical maxillary palpomere long, strongly to weakly asymmetrical ( Figs 2T, Z View Fig ); mentum ca. as long as wide, without setae along anterior margin ( Figs 2V, j–l View Fig ); pronotum 0.7–0.8× as long as wide, hood covering head forming anterior third of its length; ventral surface of the hood with set of parallel ridges ( Figs 3F, H View Fig ); prosternum without median elevation, ca. 0.3× as long as procoxal cavity ( Figs 3E, G View Fig ); procoxal cavity closed posteriorly ( Figs 3E, G, I View Fig ); mesanepisterna narrowly separated by anterior portion of mesoventrite ( E. costatus group; Fig. 3K View Fig ) or meeting mesally ( E. mendeli group; Fig. View Fig
5B) (other groups not examined); mesoventrite posteromesally with high transverse ridge ( Fig. 3K View Fig ); metaventrite ca.
as long as mesocoxa, without large smooth elevated areas ( Figs 3K View Fig , 5B View Fig ); middle and hind femora without posterior spine; phallobase short and wide; parameres simple; median lobe flat, with a pair of ventral projections or without any projections ( Figs 6L–R View Fig ); sperm pump absent; male sternite IX U-shaped ( Fig. 6K View Fig ).
Identification. The genus was revised by Pൾ*©κංඇඌ (2012) who also provided a key to the species groups and illustrated all species.
List of species (56 described species) Epimetopus plaumanni species group Epimetopus costatus species group E. multiportus Perkins, 2012 Uruguay, Paraguay (Pൾ*©κංඇඌ 2012)
E. plaumanni ( Costa Lima, 1954) Brazil (Santa Catarina, Nova Teu-
E. acuminatus Perkins, 2012 Guatemala (Pൾ*©κංඇඌ 2012) tonia) (Cඈඌඍ*ò Lංආ*ò 1954, Pൾ*©κංඇඌ E. angustus Perkins, 2012 Panama, Venezuela, Ecuador, Peru 2012)
(Pൾ*©κංඇඌ 2012) E. vianai Balfour-Browne, 1949 Argentina (B*òඅൿඈඎ*©-B*©ඈඐඇൾ 1949,
E. apocinus Perkins, 2012 Mexico, Costa Rica (Pൾ*©κංඇඌ 1979, Oඅංඏ*ò 1986, Pൾ*©κංඇඌ 2012)
2012) E. vulpinus Perkins, 2012 Brazil (Rio Grande do Sul), Uruguay
E. arizonicus Perkins, 2012 USA (Arizona) (Pൾ*©κංඇඌ 2012) (Pൾ*©κංඇඌ 2012) E. ballatoris Perkins, 2012 Venezuela, Trinidad and Tobago
(Pൾ*©κංඇඌ 2012) Epimetopus lanceolatus species group
E. bifidus Perkins, 2012 Mexico (Pൾ*©κංඇඌ 2012) E. lanceolatulus Perkins, 2012 Brazil (Mato Grosso), Paraguay E. burruyacu Oliva, 1986 Argentina (Tucuman) (Oඅංඏ*ò 1986) (Pൾ*©κංඇඌ 2012) E. costaricensis Perkins, 1979 Mexico, Belize, Costa Rica, Guate- E. lanceolatus Perkins, 2012 Brazil (Mato Grosso) (Pൾ*©κංඇඌ 2012)
mala, Honduras, Panama (Pൾ*©κංඇඌ
1979, 2012) Epimetopus trogoides species group
E. costatus ( Leconte, 1874) USA (Texas, Arkansas) (Pൾ*©κංඇඌ2012) E. clandestinus Perkins, 2012 Brazil (Mato Grosso), Venezuela E. ecuadoriensis Perkins, 2012 Ecuador (Pൾ*©κංඇඌ 2012) (Pൾ*©κංඇඌ 2012) E. fisheri Perkins, 1979 Mexico, Honduras (Pൾ*©κංඇඌ 2012), E. deceptus Perkins, 2012 Brazil (Mato Grosso) (Pൾ*©κංඇඌ 2012)
USA (Arizona) (Pൾ*©κංඇඌ 1979) E. fimbriatus Perkins, 2012 Brazil (Mato Grosso) (Pൾ*©κංඇඌ 2012)
E. hintoni Balfour-Browne, 1949 Argentina (Oඅංඏ*ò 1986, needs confir- E. tridens Perkins, 2012 Brazil (São Paulo) (Pൾ*©κංඇඌ 2012)
mation), Bolivia (B*òඅൿඈඎ*©-B*©ඈඐඇൾ E. trogoides (Sharp, 1874) Brazil (Mato Grosso, São Paulo)
1949, Pൾ*©κංඇඌ 2012) (Pൾ*©κංඇඌ 2012); records by Rඈ*ö*ü*ò
E. inaequalis Perkins, 2012 Ecuador, Peru (Pൾ*©κංඇඌ 2012) (1969) from Brazil and by Oඅංඏ*ò E. lacordairei Orchymont, 1933 Bolivia, Brazil (Mato Grosso), Pa- (1986) from Argentina need verifi-
raguay (O*©*ö*üඒආඈඇඍ 1933, Pൾ*©κංඇඌ cation
2012) E. venezuelensis Perkins, 2012 Venezuela (Pൾ*©κංඇඌ 2012)
E. latilobus Perkins, 2012 Costa Rica (Pൾ*©κංඇඌ 2012) E. latisoides Perkins, 2012 Panama (Pൾ*©κංඇඌ 2012) Epimetopus tuberculatus species group E. latus Perkins, 2012 Colombia, Venezuela (Pൾ*©κංඇඌ 2012)
E. tuberculatus Rocha, 1969 Brazil (Minas Gerais) (Rඈ*ö*ü*ò 1969,
E. lobilatus Perkins, 2012 Costa Rica (Pൾ*©κංඇඌ 2012)
Pൾ*©κංඇඌ 2012)
E. microporus Perkins, 2012 Honduras, Panama (Pൾ*©κංඇඌ 2012) E. mucronatus Perkins, 2012 Mexico, Honduras (Pൾ*©κංඇඌ 2012) E. oaxacus Perkins, 2012 Mexico (Pൾ*©κංඇඌ 2012) Records of Epimetopus E. panamensis Perkins, 1979 Panama (Pൾ*©κංඇඌ 1979, 2012) from Africa and the Arabian Peninsula E. plicatus Perkins, 2012 Venezuela (Pൾ*©κංඇඌ 2012) Specimens examined. ZAMBIA: 1 ♀ (BMNH): Kabwe, Kasanka NP, E. punctipennis Perkins, 1979 USA (Arizona, Texas, Oklahoma),
12°32ʹ28ʺS 30°12ʹ42ʺE, light trap – Edwards funnel, 30.xi.–1.xii.2012,
northern Mexico (Pൾ*©κංඇඌ1979,2012) Smith & Takano lgt. SAUDI ARABIA: 1 J (NMPC): Jizan Prov., ʻWadi
E. rectus Perkins, 2012 Costa Rica (Pൾ*©κංඇඌ 2012)
Atoudʼ, 8.ii.2016, 17°48ʹN 42°22ʹE, 245 m, J. Bezděk & D. Král lgt.
E. robustus Perkins, 2012 Panama (Pൾ*©κංඇඌ 2012) E. simplex Perkins, 1979 Costa Rica, Nicaragua, Panama, Comments. Both specimens belong to the Epimetopus
Venezuela (Pൾ*©κංඇඌ 1979, 2012)
costatus species group, i.e. the group for which the mor-
E. spatulus Perkins, 2012 Peru (Pൾ*©κංඇඌ 2012) E. steineri Perkins, 2012 Ecuador (Pൾ*©κංඇඌ 2012) phology of the male genitalia needs to be examined for E. transversoides Perkins, 2012 Peru (Pൾ*©κංඇඌ 2012) species identification. The specimen from Zambia (Fig. E. transversus Perkins, 2012 Bolivia (Pൾ*©κංඇඌ 2012) 16E) is a female and hence cannot be identified. The speci- E. trilobus Perkins, 2012 Venezuela (Pൾ*©κංඇඌ 2012) men from Saudi Arabia ( Figs 16F–G View Fig ) seems to correspond Epimetopus mendeli species group to E. burruyacu , i.e. the species endemic to Argentina. E. angulatus Balfour-Browne , Bolivia (B*òඅൿඈඎ*©-B*©ඈඐඇൾ 1949, Both specimens were collected at light. We contacted the 1949 Pൾ*©κංඇඌ 2012) collectors of both specimens who excluded the possibility E. coleuncus Perkins, 2012 Argentina, Bolivia (? Oඅංඏ*ò 1986 as of confusing or mixing the samples with those from South
E. angulatus , Pൾ*©κංඇඌ 2012) America. The specimen from Saudi Arabia is from the same
E. flavicaptus Fikáček, Barclay Ecuador (Fංκගඹൾκ et al. 2011) collecting event (same collectors, date and locality data) & Perkins, 2011 E. graniger ( Mulsant, 1851) Colombia (Pൾ*©κංඇඌ 2012) as the four specimens of the South American (likely Ar- E. mendeli Fikáček, Barclay Peru (Fංκගඹൾκ et al. 2011, Pൾ*©κංඇඌ gentinian) Chaetarthria reported by Fංκගඹൾκ & Lංඎ (2019: & Perkins, 2011 2012) 251). Most likely, these specimens have been mislabeled. E. peruvianus Perkins, 2012 Peru (Pൾ*©κංඇඌ 2012) E. thermarum species group Discussion E. arcuatus Perkins, 2012 Paraguay (Pൾ*©κංඇඌ 2012) The aim of this study was to publish the newly accu- E. balfourbrownei Rocha, 1969 Brazil (Mato Grosso) (Rඈ*ö*ü*ò 1969, mulated data on the family Epimetopidae which became
Pൾ*©κංඇඌ 2012)
available due to the newly collected material. Fresh alcohol
E. clypeatus Perkins, 2012 Guyana (Pൾ*©κංඇඌ 2012) E. surinamensis Perkins, 2012 Suriname (Pൾ*©κංඇඌ 2012) specimens enabled us to provide the first DNA sequences E. thermarum Schwarz & USA (Arizona, Texas), Mexico, of Eumetopus and Eupotemus . The new material from Barber, 1917 Belize, Guatemala, Costa Rica, Africa made it possible to dissect some specimens and
Panama, Venezuela, (S*ö*üඐ*ò*©ඓ & perform morphological comparative studies based on
B*ò*©*ôൾ*© 1918, Rඈ*ö*ü*ò 1969, Pൾ*©κංඇඌ
all three genera. New records complementing the data
2012)
on the distribution of all three genera became available. the air bubble is usually partly formed with the help of the Yet, it is very clear that the knowledge about the family antenna (H*©*ôගඹൾκ 1950), and the antennal modifications remains rather limited in some aspects; these are defined in Georissidae and Epimetopidae may hence correspond
and discussed below. to the adapted way of the gas exchange not necessarily
indicating a close relationship of both families.
Phylogenetic position of the family. It was mention-
Larvae. Larvae are so far only known for a few species
ed above that there is an apparent and strong conflict
of the genus Epimetopus (Rඈ*ö*ü*ò 1967, 1969; Cඈඌඍ*ò et between the position of the Epimetopidae revealed by
al. 1988; A*©*ö*ü*òඇǤൾඅඌκඒ 1997; Fංκගඹൾκet al. 2011; Rඈmorphological and molecular characters. Analyses based
ൽ*©ංǤඎൾඓ et al. 2020) but unknown for the other two genera.
on morphology always place Epimetopidae as sister to
Epimetopus larvae are all characterized by the adaptations Georissidae , irrespectively of what kind of characters are
for the underwater processing of the prey by piercing and used, and whether adult or larval data are included (H*òඇඌൾඇ
sucking: the adapted form of the mandibles, the enlarge- 1991; Bൾඎඍൾඅ 1994, 1999; A*©*ö*ü*òඇǤൾඅඌκඒ 1998; Bൾඎඍൾඅ
ment of the epistomal lobes and the reduction of the labrum
& Kඈආ*ò*©ൾκ 2004; Bൾඎඍൾඅ & Lൾඌ*ö*üൾඇ 2005; Bൾ*©ඇ*ü*ò*©ൽ
(Rඈൽ*©ංǤඎൾඓ et al. 2020). They are often associated with
et al. 2009; Fංκගඹൾκ et al. 2012). In contrast, molecular
reductions of spiracles and the closure of the tracheal analyses, despite not being conclusive about the phylo-
system (Rඈൽ*©ංǤඎൾඓ et al. 2020). Similar morphology of genetic position of the Epimetopidae never place them
the head and mouthparts, associated with underwater prey close to Georissidae (Bൾ*©ඇ*ü*ò*©ൽ et al. 2006, 2009; S*üඈ*©ඍ
processing evolved independently in Epimetopidae and in
& Fංκගඹൾκ 2013; M*öKൾඇඇ*ò et al. 2014; Lඳ et al. 2020).
three unrelated groups of the Hydrophilidae (Fංκගඹൾκ et
If the molecular data are correct, it would imply that the
al. 2018, Rඈൽ*©ංǤඎൾඓ et al. 2020). Moreover, Rඈൽ*©ංǤඎൾඓ supposed synapomorphies of Epimetopidae + Georissidae
et al. (2020) noticed that the lineages sister to those with evolved in both groups independently, as a result of con-
piercing-sucking mouthparts often have very different vergent evolution. We document here that Epimetopidae
morphology of the head and a tracheal system well correinhabit moist sandy shores of streams of standing waters,
sponding to the usual hydrophilid morphology. In addition,
i.e. the same environment as most Georissidae (Mൾඌඌඇൾ*©
at least in two cases in the Hydrophilidae ( Laccobiini and 1965, 1972; Fංκගඹൾκ & F*òඅ*òආ*ò*©ඓං 2010; Lංඍඈඏκංඇ &
Berosini ), the lineage with piercing-sucking larval adaptati- Fංκගඹൾκ 2011; Lංඍඈඏκංඇ 2018) and some riparian groups
ons contains significantly more species than its sister clade
of Hydrophilidae (e.g., Chaetarthria Stephens, 1835 and
in which larvae process the food above the water. This Thysanarthria Orchymont, 1926 : Pൾ*©κංඇඌ 1976, Fංκගඹൾκ
observation resembles the situation in the Epimetopidae .
& Lංඎ 2019). Chaetarthria and Thysanarthria are deeply
The species-rich Epimetopus with 56 species has larvae nested clades of the Hydrophilidae (S*üඈ*©ඍ & Fංκගඹൾκ
with piercing-sucking mouthparts. Its sister Eupotemus 2013 ) and are not closely related to Epimetopidae . Still,
has only eight known species and the larvae are unknown.
they bear some of the characters considered as synapo-
Hence, we cannot exclude that larvae of Eupotemus and morphies of Georissidae + Epimetopidae : they have a very
Eumetopus may be not adapted for piercing-sucking food long antennal scape, a bulbose pedicel, strongly reduced
processing, and hence may look different from those of
(yet not totally absent) pubescence on the ventral body
Epimetopus in head and mouthpart morphology and in surface, reduced gula and hence fused gular sutures, and
the development of the larval tracheal system. The first they bear numerous digitiform sensilla on the base of the
instar larvae of Eumetopus and Eupotemus can be obtained maxillary palpomere IV (Fංκගඹൾκ & Lංඎ 2019). Moreover,
from the egg cases carried by the females and should be
the mentum of Thysanarthria bears series of long setae
studied in detail.
along its anterior margin (Fංκගඹൾκ & Lංඎ 2019: fig. 3A),
Egg cases carried by females. Egg cases are carried by strongly resembling the situation found in Eumetopus (Fig.
females of all three epimetopid genera, and hence represent 4D). These convergences with Hydrophilidae indicate that
a synapomorphy of Epimetopidae . Similar behavior is
the convergent evolution of these characters cannot be a
present in two unrelated lineages of Hydrophiloidea : the priori excluded for Georissidae and Epimetopidae . Addi-
Spercheidae (Fංκගඹൾκ 2019d) and the Helochares group tional studies on the biology and functional morphology
of the hydrophilid subfamily Acidocerinae (S*üඈ*©ඍ et al.
of both latter families are needed to understand whether
2021). The egg-carrying behavior is considered as derived their biology and the morpho-functional adaptations to
in the Hydrophiloidea (H*òඇඌൾඇ 2000), i.e., it evolved indedeal with the riparian environment are indeed identical
pendently in each mentioned lineage and may be adaptive.
or just analogous. For example, the pronotal hood is a
This seems to be corroborated by the slightly different way unique character shared by both families. The studies of
in which the egg cases are carried in each group (H*òඇඌൾඇ the function of the hood including the parallel ridges on
2000). The purpose of this adaptation and whether similar
its ventral side (see Biology of Epimetopidae above for
or clade-specific evolutionary pressures led to the evolution current hypotheses) and a detailed comparative study of
of this behavior remains to be tested.
these structures between Georissidae and Epimetopidae
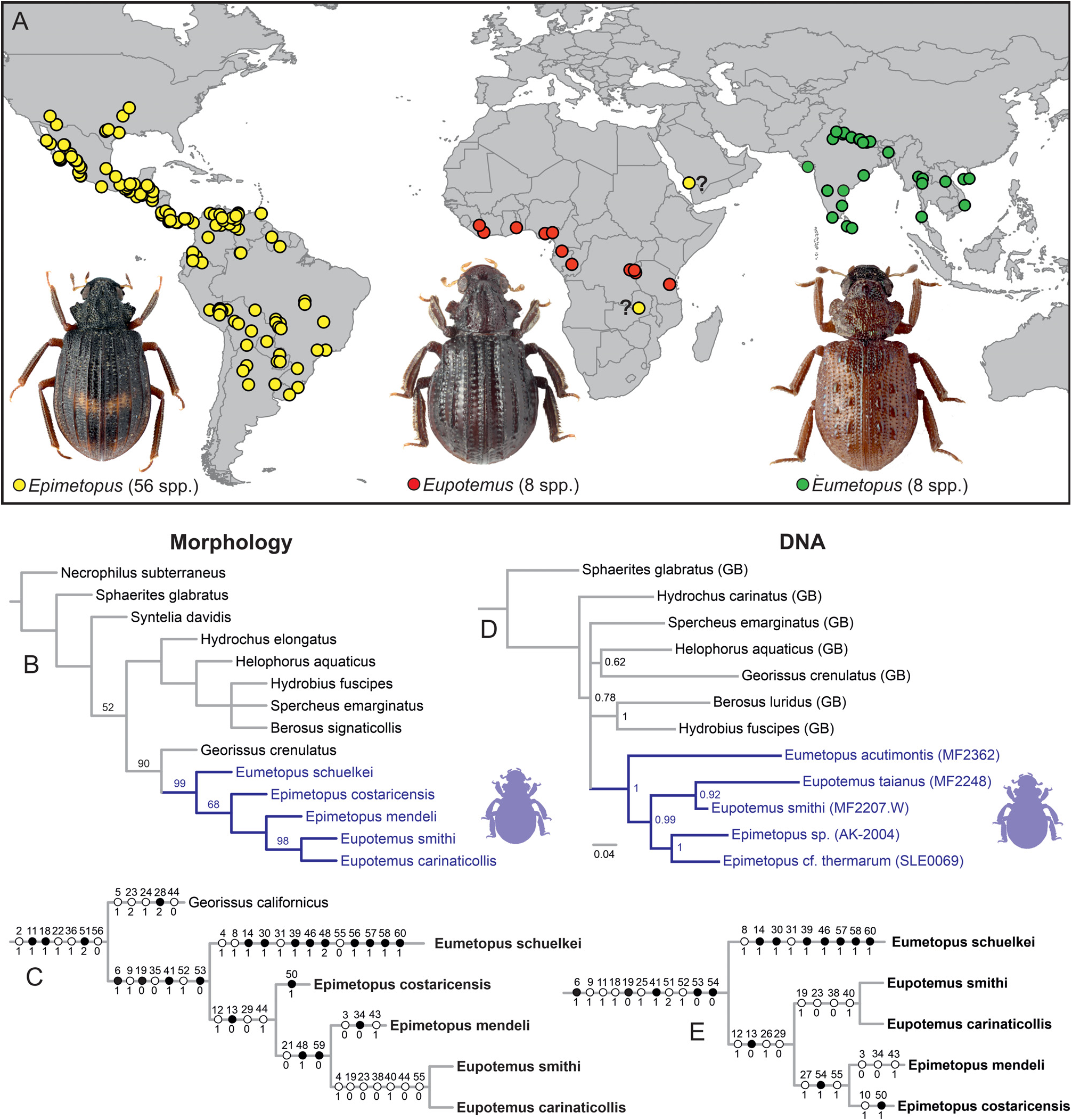
Monophyly and internal topology of Epimetopus . Our may reveal useful information. The gas exchange is ano-
morphological analysis failed to reveal the monophyly of
ther unusual aspect which is shared by Georissidae and
the genus Epimetopus ( Figs 1B–C View Fig ). In contrast, the DNA- Epimetopidae : both clades lack the hydrofuge pubescen-
-based analysis indicated Epimetopus as monophyletic.
ce, which, in other Hydrophiloidea , holds the ventral air
The taxon sampling was different in both analyses. Hence, bubble (Fංκගඹൾκ 2019c, this paper). In Hydrophiloidea ,
both topologies may not be incongruent: the paraphyly of Epimetopus in the morphology analysis is caused by the member of the E. costatus group which is likely not involved in the molecular analysis. All Epimetopus species have posteriorly closed procoxal cavities, unlike any other epimetopids, based on which we consider the paraphyly of Epimetopus as unlikely. However, we cannot totally exclude it based on our data. Epimetopus is morphologically much more diverse than Eumetopus and Eupotemus .
This is evident even from our limited taxon sampling containing two Epimetopus species , i.e. representatives of the E. costatus and E. mendeli groups. The comparison of these two species revealed numerous differences, e.g., in the form of the prothoracic hypomeron (compare Figs 3E and G View Fig ), in the form and sculpture of the meso- and metaventrite (compare Figs 3K View Fig and 5B View Fig , for additional SEMs of E. mendeli see also Fංκගඹൾκ et al. 2011), in the form of the trochanters (with dorsal plates in E. costatus group, without such plates in E. mendeli ) and in the tarsal formula (5-5- 5 in E. mendeli , 4-4- 4 in E. costatus group).
These differences indicate that the E. costatus group may indeed have a rather isolated position within Epimetopus ; that needs to be tested by analyses with a wider species sampling covering all species groups of Epimetopus . Some other species groups also show apparent differences, e.g., in the eye morphology (completely divided in E. trogoides group, partially divided in other species; compare Figs 11 View Fig and 12 View Fig in Fංκගඹൾκ et al. 2011) and in the morphology of the male genitalia (with ventral projections likely corresponding to those of Eupotemus or without such projections; Figs 6L–R View Fig and genitalia illustrations in Pൾ*©κංඇඌ 2012). Additional studies are needed to understand the evolution of these characters within the genus and to confirm the monophyly of Epimetopus .




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Epimetopus Lacordaire, 1854
| Fikáček, Martin, Matsumoto, Keita, Perkins, Philip, Prokin, Alexander, Sazhnev, Alexey, Litovkin, Stanislav & Jäch, Manfred A. 2021 |
Sepidulum
| LECONTE J. L. 1874: 47 |
Epimetopus
| LACORDAIRE T. 1854: 467 |
Ceratoderus
| MULSANT E. 1851: 1 |