
Terebellides gracilis (Malm, 1874)
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3691.3.3 |
|
publication LSID |
lsid:zoobank.org:pub:24C5A895-F6B0-47CF-B138-A74670268984 |
|
DOI |
https://doi.org/10.5281/zenodo.6164482 |
|
persistent identifier |
https://treatment.plazi.org/id/038B0C19-FFFF-FF8B-3FC4-B4340A7CFC0C |
|
treatment provided by |
Plazi |
|
scientific name |
Terebellides gracilis (Malm, 1874) |
| status |
|
Terebellides gracilis (Malm, 1874) View in CoL
( Figures 2–4 View FIGURE 2 View FIGURE 3 View FIGURE 4 , 12 View FIGURE 12 b, 13)
Terebellides gracilis Malm 1874: 100 .
Terebellides gracilis —Hansson 1998: 77. Parapar et al. 2011: 11, Figs 8–10 View FIGURE 8 View FIGURE 9 View FIGURE 10 , 13 View FIGURE 13 c. Terebellides williamsae —Jirkov 1989: 124. Jirkov 2001: 529, Figs 1–4 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 .
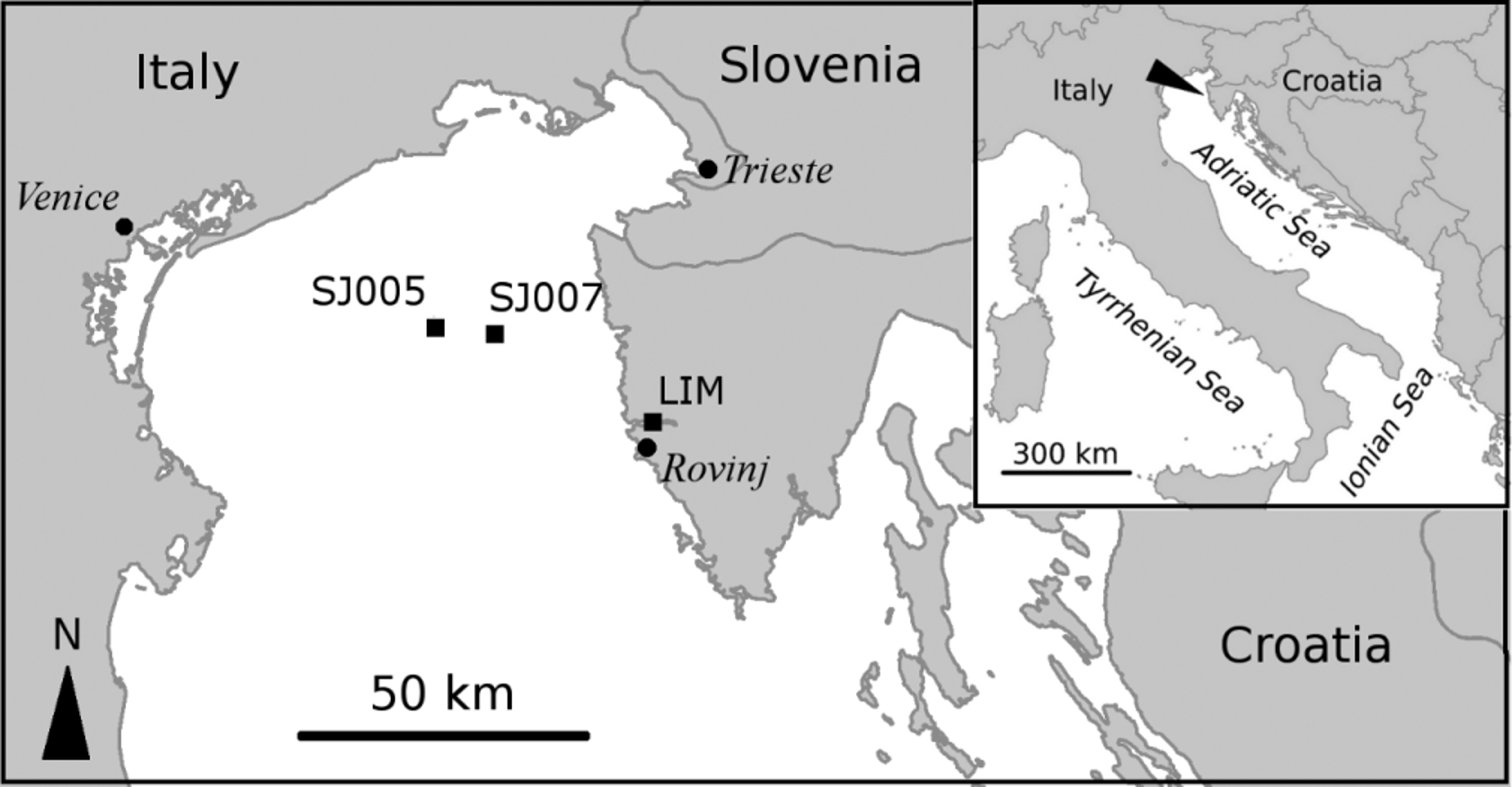
Material examined. A total of 23 specimens were examined (27.4% of the total Terebellides specimens collected) all from station SJ 0 0 7: 27.02.2003 (PMR-14550, 2 specs.; PMR-14551, 1 spec.; PMR-14552, 1 spec. on SEM stub; PMR-14553, 1 spec. on SEM stub); 28.05.2003 (PMR-14554, 2 specs.; PMR-14555, 3 specs); 12.08.2003 (MNCN 16.01/14714, 1 spec.; MNCN 16.01/14715, 2 specs.); 23.01.0 4 (2 specs.) (coll. BM); 30.08.2004 (2 specs.) (coll. BM); 0 2.12.2004 (PMR-14556, 3 specs.; PMR-14557, 3 specs.).
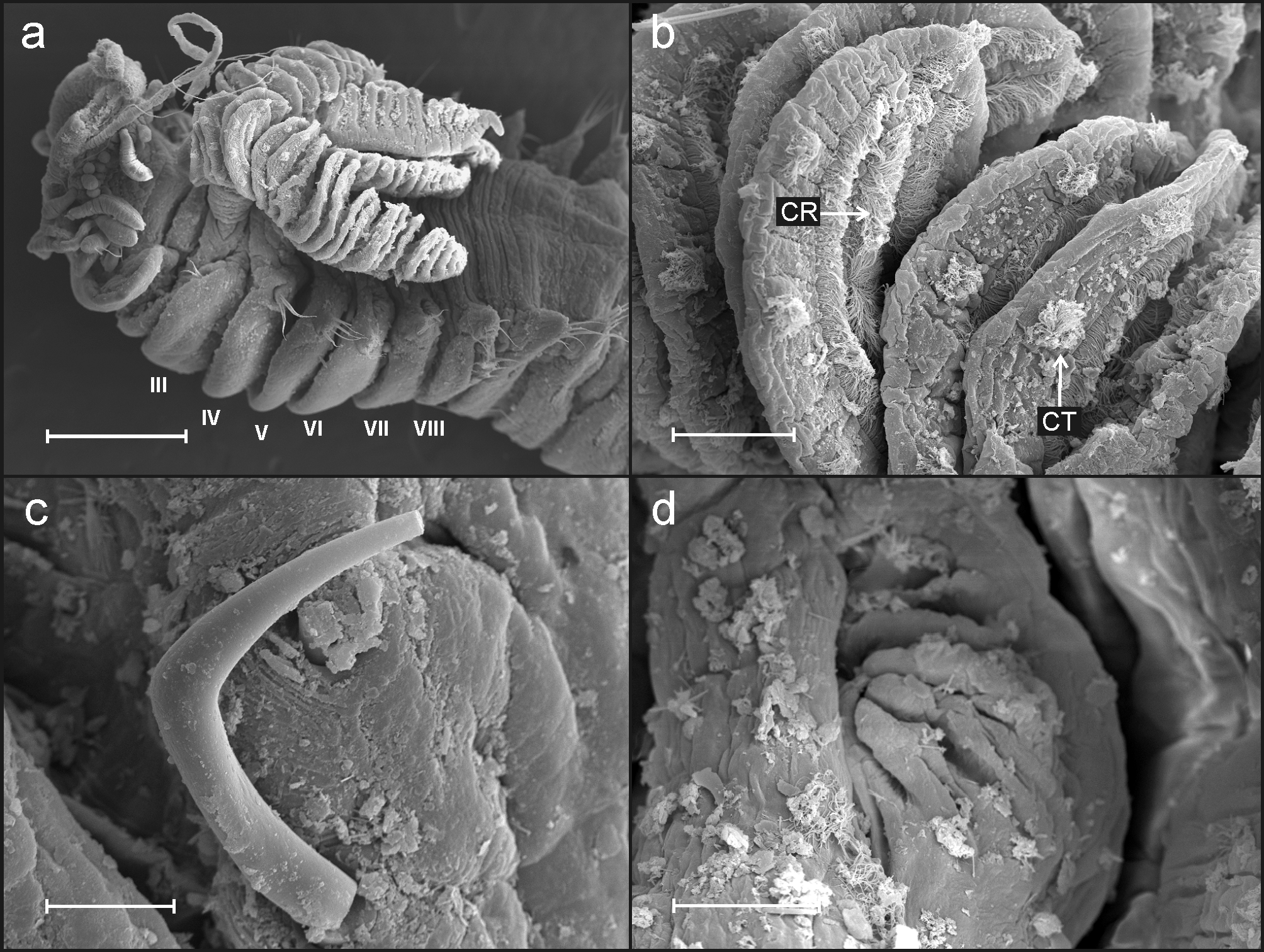
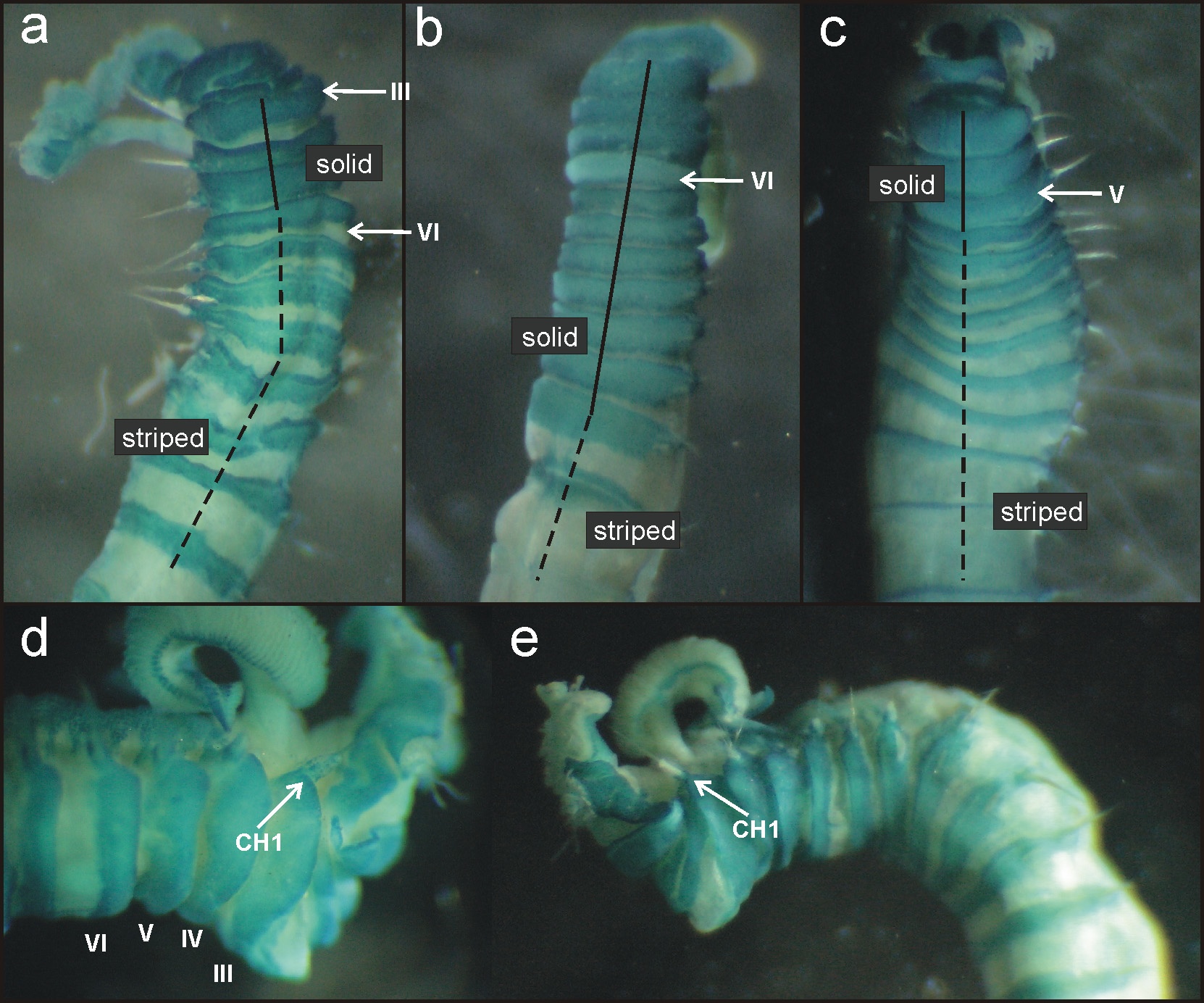
Description of Adriatic Sea specimens. Complete specimens range from 12 to 18 mm in length and 1.2 to 0.7 mm in width; body tapering posteriorly with segments becoming increasingly shorter and crowded towards pygidium. Prostomium compact; tentacular membrane surrounding mouth, with many long buccal tentacles with expanded tips. SGI forming an expanded structure below tentacular membrane. Lateral lappets on SGIII–VIII ( CH 1–6) ( Fig. 2 View FIGURE 2 a) being larger and continuing ventrally on SG III to VI, declining in size posteriorly. No conspicuous dorsal rounded projection in anterior chaetigers or oval-shaped glandular region in CH 3.
Branchiae arising as single structure from SGIII, consisting of single stalked structure located mid-dorsally ( Figs 2 View FIGURE 2 a, 3a) made up of two posterior pairs of lobes fused along less than one third of their length; lower pair slightly shorter than upper pair. Pointed projection of posterior region of lobes present in lower lobes. Anterior branchial projection (fifth lobe) not conspicuous. Both sides of branchial lamellae provided with several parallel bent rows of cilia and outer tufts of cilia ( Figs 2 View FIGURE 2 b, c; 3b). Branchial lamellae less tightly packed than in the other two Adriatic species.
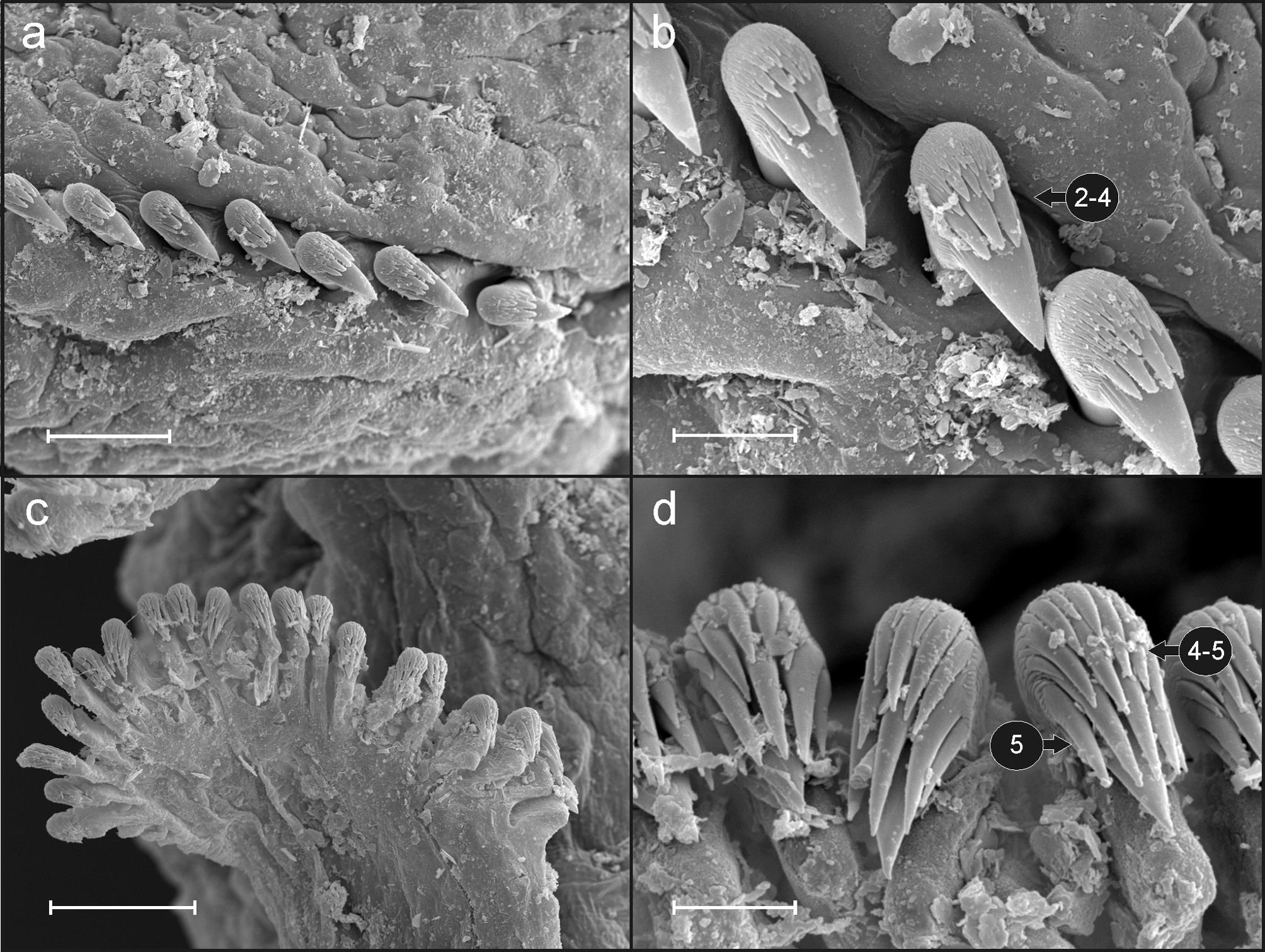
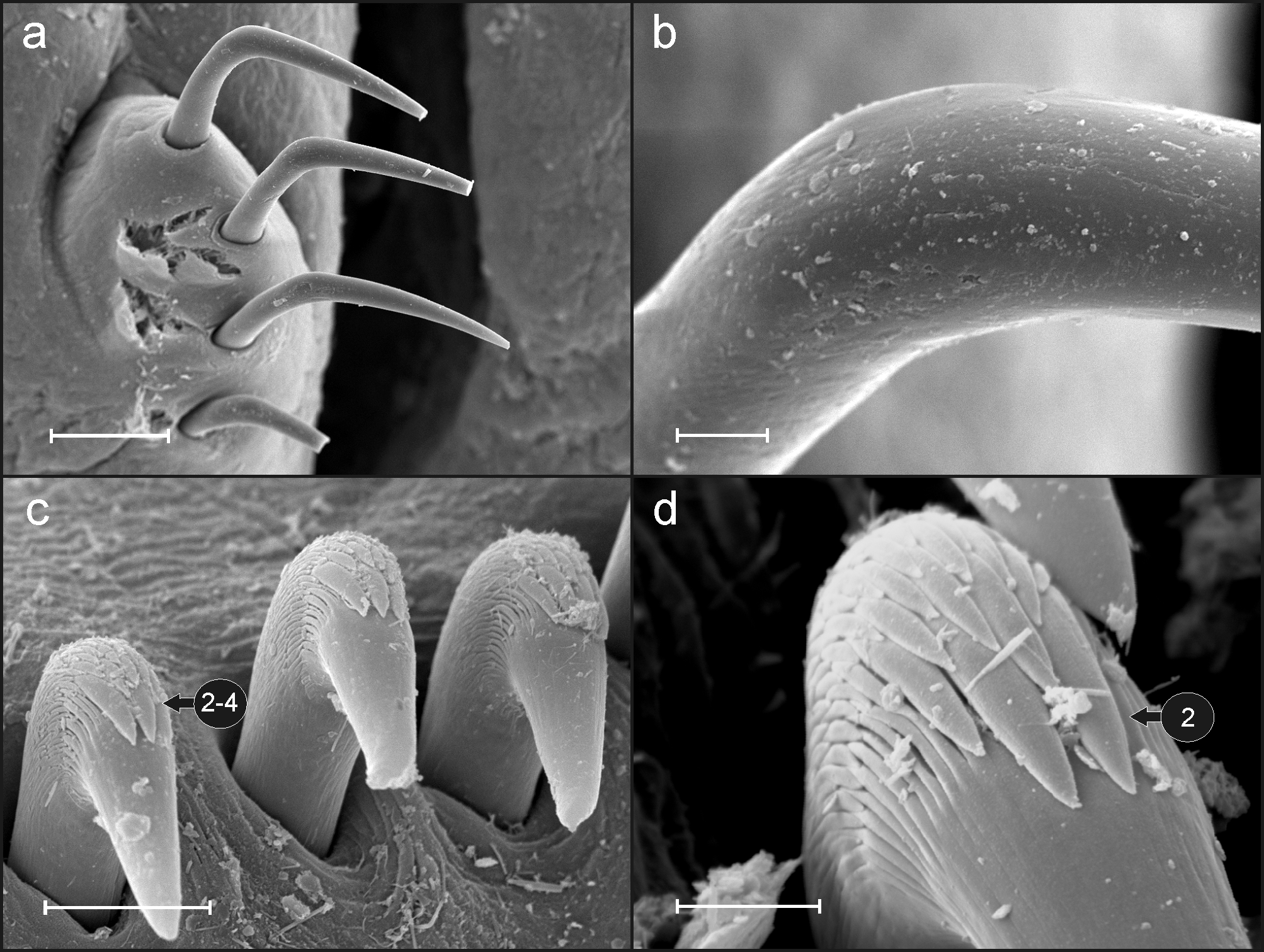
Eighteen pairs of thoracic notopodia (SGIII–XX). Notopodia and notochaetae of first chaetiger not visible in some specimens ( Fig. 2 View FIGURE 2 a, d); chaetae present only in larger specimens but much shorter than on subsequent notopodia ( Fig. 3 View FIGURE 3 a). Neuropodia present as sessile pinnules from CH 6 (SGVIII) to pygidium and provided with uncini in single rows throughout ( Fig. 4 View FIGURE 4 a). The absence of notochaetae in first chaetiger in most specimens may lead to the erroneous observation that first neuropodium is located on CH 5 instead of CH 6. First thoracic neuropodia provided with 5 – 7 sharply bent, acute tipped, geniculate acicular hooks ( Fig. 3 View FIGURE 3 c). Minute teeth forming a capitium on geniculate chaetae not observed. Second and all subsequent thoracic neuropodia with up to 7–10 uncini per torus ( Fig. 4 View FIGURE 4 a). Uncini provided with long shafted denticulate hooks with 2 – 4 teeth above main fang surmounted by 4 – 5 shorter teeth and an upper crest of several smaller denticles ( Fig. 4 View FIGURE 4 b), dental formula MF:2– 4:4–5:∞.
Between 30−32 abdominal neuropodia as erect pinnules provided with about 22 uncini per torus ( Fig. 4 View FIGURE 4 c); uncini with 5 teeth above main fang surmounted by a crest of 4–5 shorter teeth and a variable number of smaller teeth ( Fig. 4 View FIGURE 4 d); dental formula MF:5:4–5:∞. Pygidium blunt, funnel-like depression.
One large, button-hole like nephridial opening on each notopodium of CH 4 and CH 5 ( Fig. 3 View FIGURE 3 d).
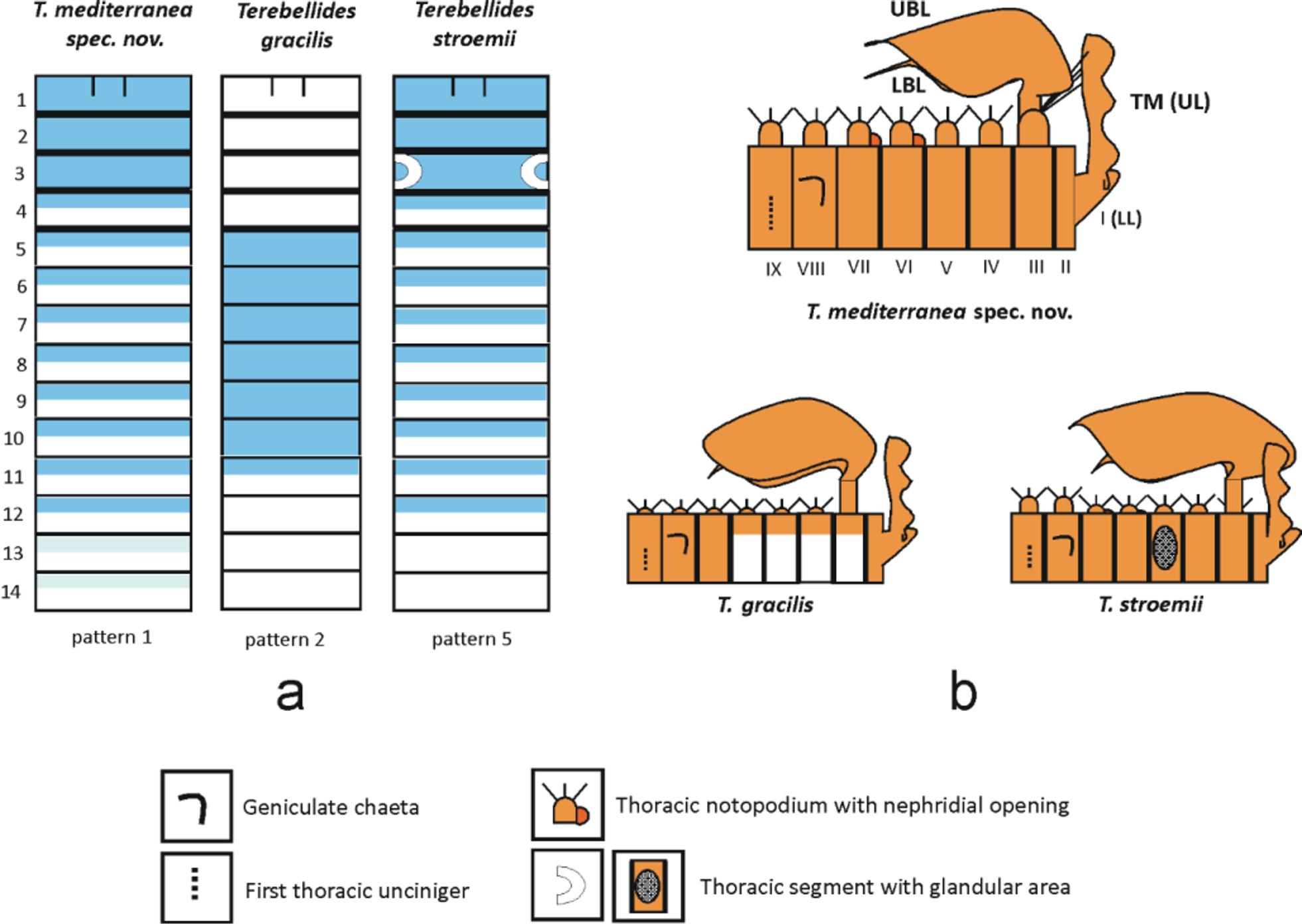
MG staining pattern 2 ( Figs 12 View FIGURE 12 b, 13a): compact green colouration in CH 1 – 10, quickly fading in following segments. CH 1 – 3 slightly and CH 4 much less stained than anterior and posterior ones. Colour in alcohol pale brown with four anterior thoracic chaetigers ventrally white ( Fig. 13 View FIGURE 13 b).
Remarks. Hansson (1998) proposed to recover T. gracilis from the synonymy of T. stroemii due to the presence of four white ventrally colored thoracic segments (SGIII–VI; CH 1–4) in T. gracilis . Following this opinion, Parapar et al. (2011) redescribed the species from material collected in the BIOICE project in Iceland and proposed also T. williamsae Jirkov, 1989 as a junior synonym.
Terebellides gracilis was until now found in the Norwegian and Barents Seas (Malm 1874; Jirkov 1989) and Iceland (Parapar et al. 2011). Our Mediterranean records extend the range of the species into quite different ecological conditions compared to those where it was recorded so far. However, the finding of this species in the Northern Adriatic Sea is not so surprising and it supports the strong boreal affinity of this region, as well as its ecological and biogeographical similarities with the North Atlantic, which have previously been documented (Bianchi et al. 2004; Boero & Bonsdorff 2007). The Adriatic Sea is the northernmost Mediterranean region and together with the Gulf of Lions and the Northern Aegean Sea the coldest sector of the Mediterranean. It is geomorphologically, hydrographically and biogeographically a peculiar Mediterranean region with lower average temperatures that allow the presence of a specific flora and fauna with cold water affinities (Mikac & Musco 2010). Our research showed that Adriatic specimens of T. gracilis are slightly shorter than boreo-arctic specimens. This feature could be a morphological variability due to the different ecological conditions in which specimens are living. Further molecular analyses could possibly assist to clarify this phenomenon.
Adriatic specimens are slightly shorter (12–18 vs 10–25 mm) and have less abdominal chaetigers (30–32 vs. about 43) and less uncini (22 vs. 45) than boreo-arctic specimens. Also, the chaetae from first thoracic chaetiger are less conspicuous than in Icelandic specimens and the dental formula of thoracic uncini differs, with two teeth surmounting the main fang in North Atlantic specimens (MF:2:5:∞) but 2−4 in Adriatic ones (MF:2−4:4−5:∞).
The staining pattern coincides with one of the Icelandic specimens although Parapar et al. (2011) erroneously reported pattern 4 for those specimens instead of 2 (Parapar et al. 2011, page 19, Table 1).
Habitat. Found at shelf depths (385–390 m) in Scandinavian and Russian waters (Jirkov, 1989; 2001; Hansson, 1998). North and South coast of Iceland; 68 to 2076 meters depth (Parapar et al., 2011). In the Adriatic Sea the species was found on the offshore station in silty sand at 31 m.
Distribution. Norwegian Sea and Barents Sea (Malm 1874; Jirkov 1989); Iceland (Parapar et al. 2011) and Adriatic Sea. Our finding in the Adriatic Sea represents the first record of this species in the Mediterranean Sea, and considerably expands the southern boundary of this species.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |