
Terebellides mediterranea, Parapar, Julio, Mikac, Barbara & Fiege, Dieter, 2013
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3691.3.3 |
|
publication LSID |
lsid:zoobank.org:pub:24C5A895-F6B0-47CF-B138-A74670268984 |
|
DOI |
https://doi.org/10.5281/zenodo.6164484 |
|
persistent identifier |
https://treatment.plazi.org/id/038B0C19-FFF8-FF87-3FC4-B24E0ACAFED7 |
|
treatment provided by |
Plazi |
|
scientific name |
Terebellides mediterranea |
| status |
sp. nov. |
Terebellides mediterranea View in CoL spec. nov.
( Figures 5–8 View FIGURE 5 View FIGURE 6 View FIGURE 7 View FIGURE 8 , 12 View FIGURE 12 a, d, e, 13; Table 1)
Material examined. A total of 10 specimens were examined (11.9% of the total Terebellides specimens collected). Type material: Station SJ 0 0 5 - 30.08.2004 (PMR-14558, 2 paratypes). Station SJ 0 0 7 - 27.02.2003 (PMR- 14559, 1 holotype; PMR-14560, 1 paratype on SEM stub); 28.05.2003 (PMR-14561, 1 paratype; PMR-14562, 1 paratype on SEM stub); 29.01.2004 (MNCN 16.01/14716, 1 paratype).
Non-type material: Station SJ 0 0 5 - 27.02.2003 (1 spec.) (coll. BM). SJ 0 0 5 - 30.08.2004 (2 specs.) (coll. BM).
Comparative material: Terebellides californica Williams, 1984 . Paratype AM W197111.
Type specimens of T. californica were requested to the Los Angeles California Natural History Museum, but unfortunately they were not available for study. However, general characteristics of this species presented in Hutchings and Peart (2000) and Schüller and Hutchings (2010) were corroborated by P. Hutchings who kindly checked the paratype of this species deposited in the Australian Museum (Sydney) (AM).
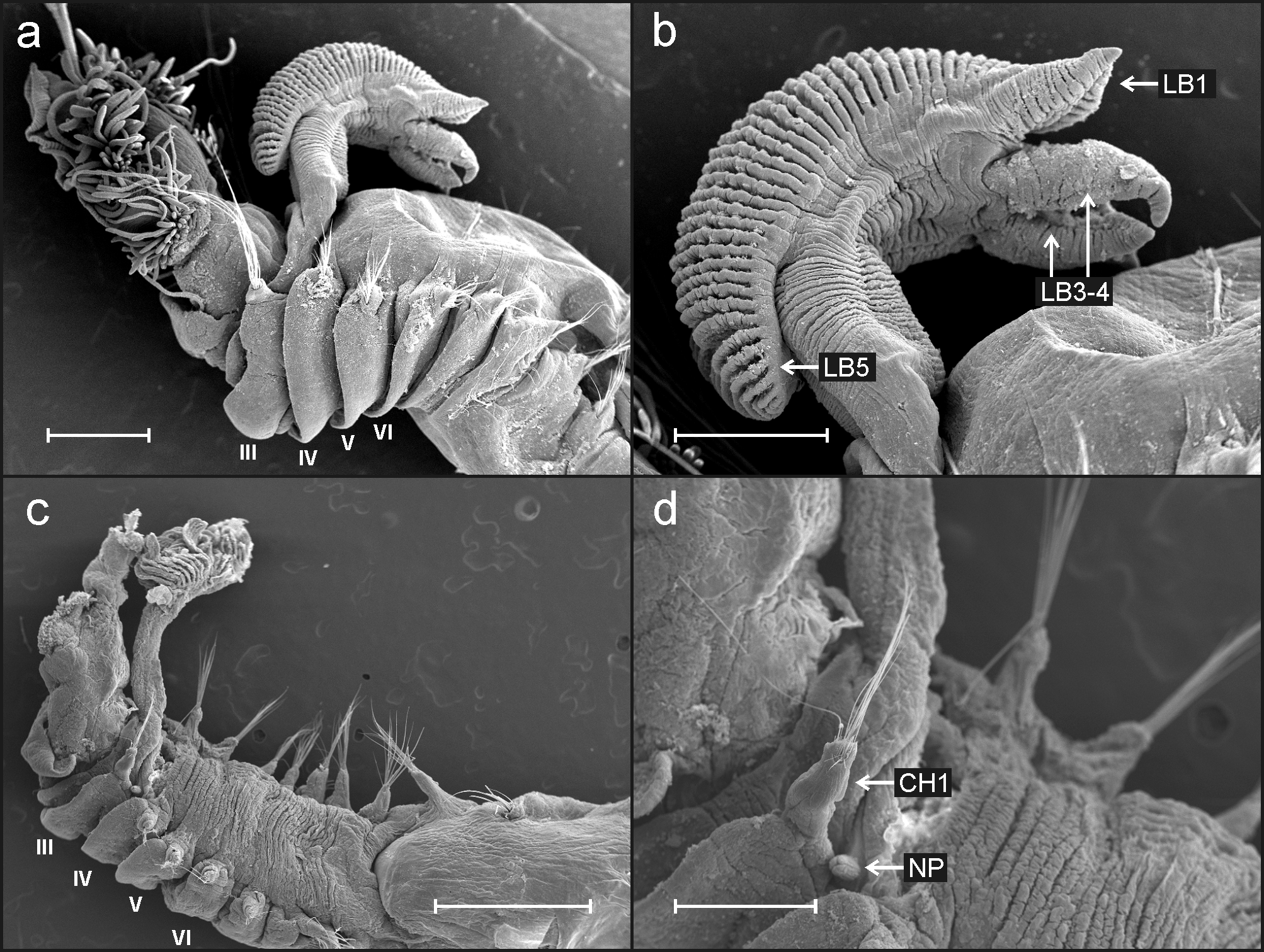
Description based on holotype. Complete specimen, 23 mm long and 2.0 mm wide; body tapering posteriorly with segments increasingly shorter and crowded towards pygidium. Prostomium compact; large tentacular membrane surrounding the mouth provided with many long buccal tentacles with expanded tips ( Fig. 5 View FIGURE 5 a, 8a). SGI forming an expanded structure below tentacular membrane. Lateral lappets on SGIII–VII, CH 1–5, being larger and continuing ventrally in SGIII–V declining in size posteriorly ( Fig. 5 View FIGURE 5 a). No conspicuous dorsal rounded projection on anterior chaetigers or oval-shaped glandular region in CH 3.
Branchiae arising as single structure from SGIII, consisting of a single stalked structure situated mid-dorsally ( Fig. 5 View FIGURE 5 a) made up of two pairs of lobes fused along most of their lengths; lower pair much shorter then upper pair. Pointed projection of posterior region of both upper and lower lobes and large anterior branchial projection (fifth lobe) present ( Fig. 5 View FIGURE 5 b). Both sides of branchial lamellae provided with several parallel bent rows of cilia ( Fig. 6 View FIGURE 6 a) and tufts of cilia on the outer edge ( Fig. 6 View FIGURE 6 b).
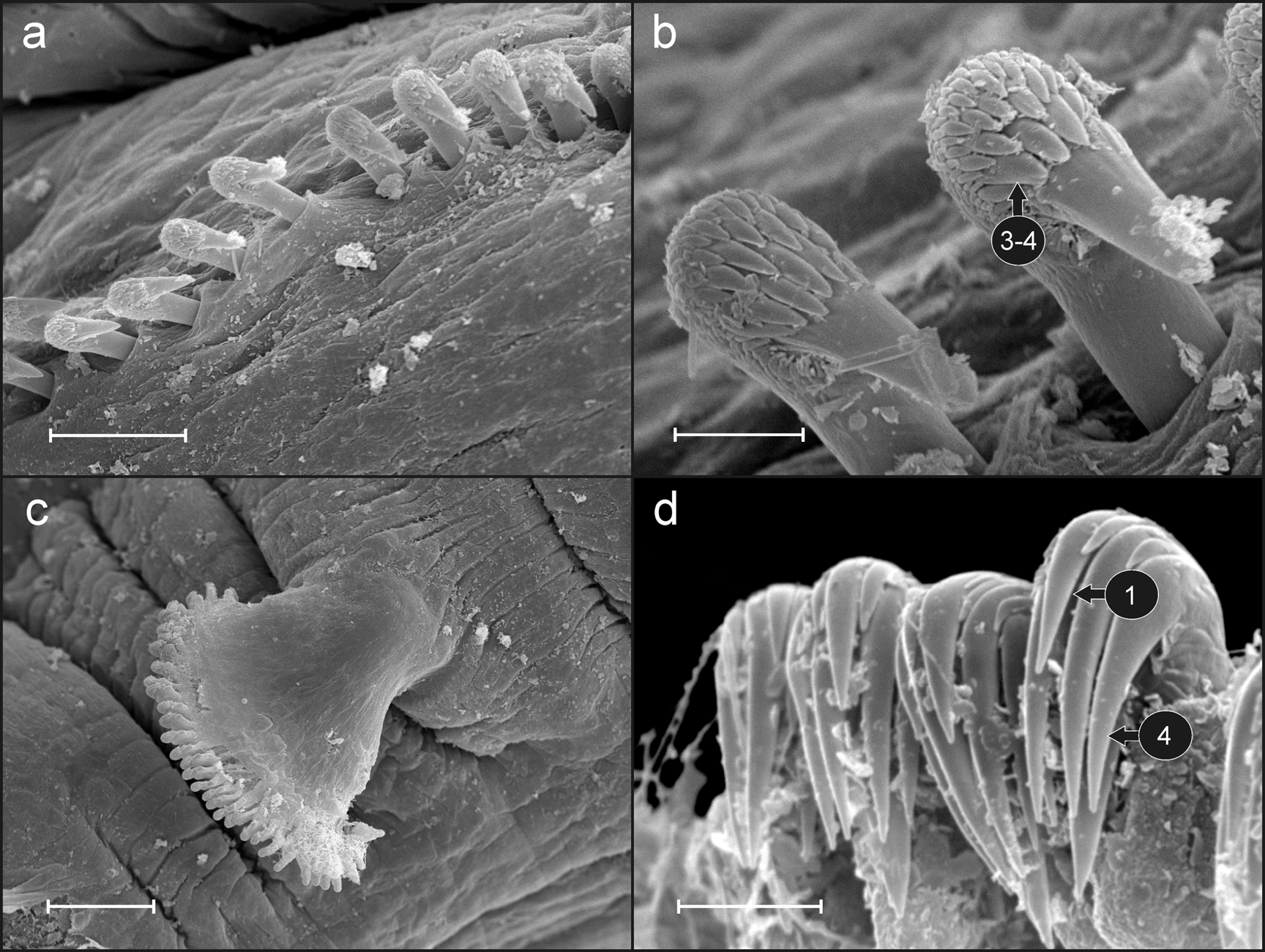
Eighteen pairs of thoracic notopodia (SGIII −XX). Notopodia present from CH 1, larger than subsequent notopodia ( Fig. 5 View FIGURE 5 a, c, d); notochaetae of CH 1 much longer than following notochaetae. All notochaetae simple capillaries with textured surface ( Fig. 6 View FIGURE 6 c). Neuropodia present as sessile pinnules from CH 6 (SGVIII) to pygidium and provided with uncini in single rows starting from CH 7 (SGIX) throughout. First thoracic neuropodia ( CH 6) provided with 4–5 sharply bent, acute tipped, geniculate acicular hooks ( Fig. 6 View FIGURE 6 d). Minute teeth forming a capitium on geniculate chaetae not observed. Second and all subsequent thoracic neuropodia with up to 10–16 uncini per torus ( Fig. 7 View FIGURE 7 a). Uncini provided with long shafted denticulate hooks with 3−4 teeth above main fang surmounted by 6−7 shorter teeth and an upper crest of several smaller denticles ( Fig. 7 View FIGURE 7 b), dental formula MF:3–4:6–7:∞. Thirty-two abdominal neuropodia as erect pinnules provided with about 33 uncini per torus ( Fig. 7 View FIGURE 7 c); uncini with four teeth above main fang surmounted by a fifth tooth in the middle, an upper crest of 4 teeth ( Fig. 7 View FIGURE 7 d) and a variable number of smaller teeth, dental formula MF:4:1:4:∞.
One large papilla located behind first thoracic notopodia ( Figs 5 View FIGURE 5 d; 8b), and two button-hole like pairs of nephridial openings; located dorsal to each notopodium of SGVI and VII ( CH 4 and 5) ( Fig. 8 View FIGURE 8 c, d). Pygidium blunt, funnel-like depression.
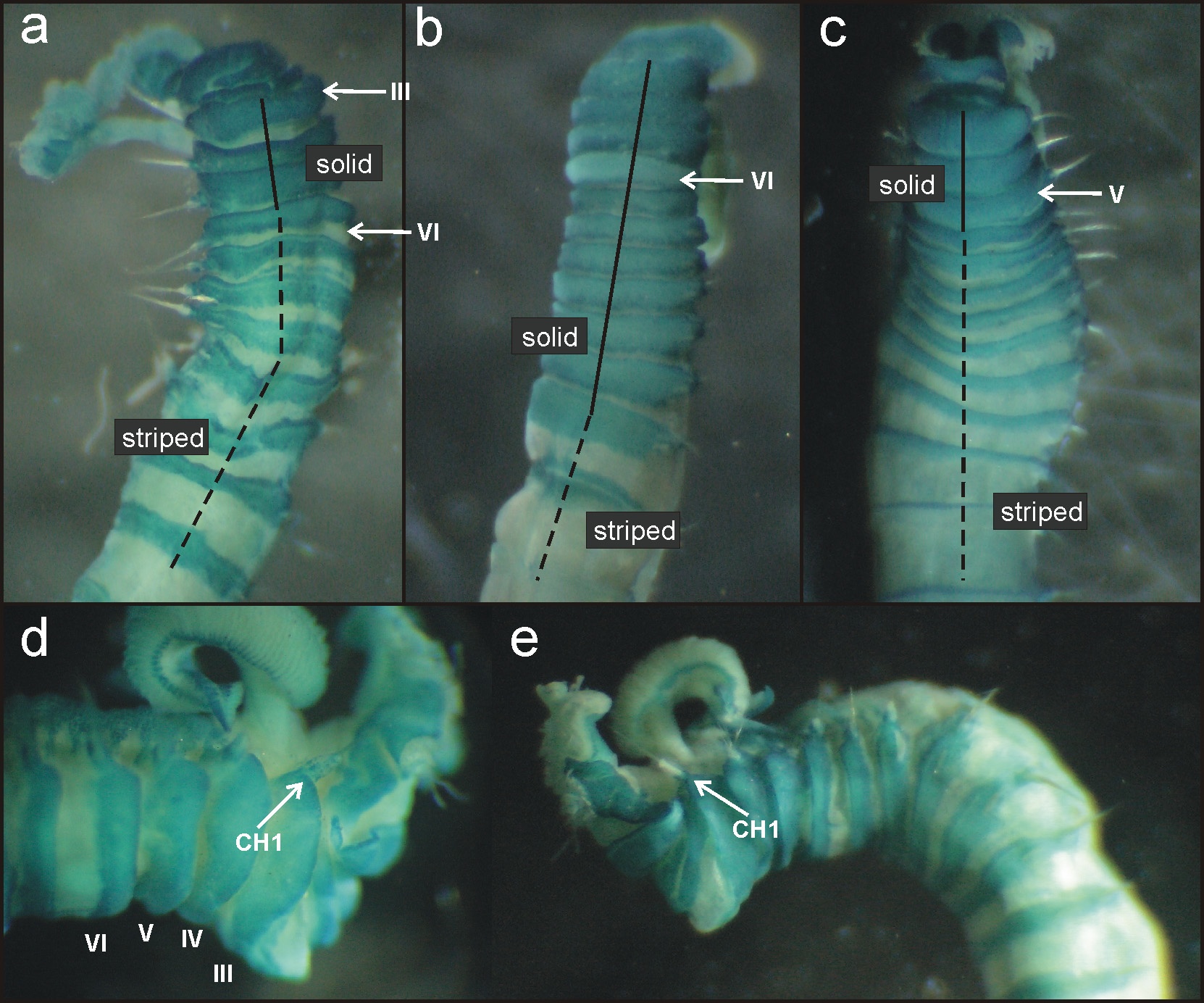
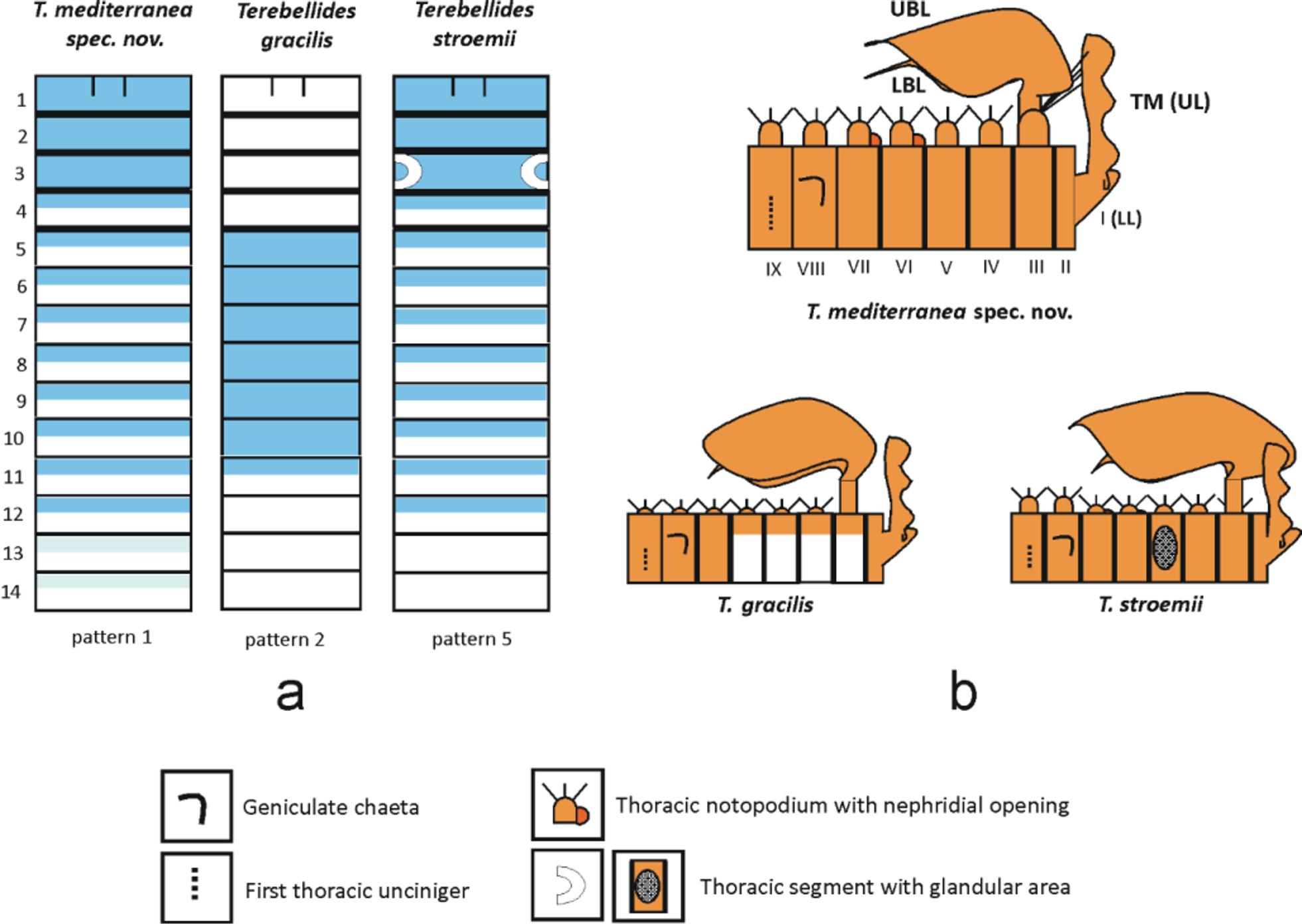
MG staining pattern 1 ( Figs 12 View FIGURE 12 a,d,e; 13a): compact green colouration in CH 1 – 3, then turning into striped pattern in CH 4 – 12 and fading in the following segments. Colour in alcohol pale brown ( Fig. 13 View FIGURE 13 b).
Remarks. The presence of large notopodia provided with long notochaetae in the first thoracic chaetiger is a character usually employed to discriminate species in taxonomic keys of the genus Terebellides (Garraffoni & Lana 2003; Solis-Weiss et al. 2009) and also in phylogenetic studies (Garraffoni & Lana 2004). Following Hutchings and Peart (2000) two species have this diagnostic character: T. kobei Hessle, 1917 and T. californica Williams, 1984 .
Terebellides kobei was described by Hessle (1917) from specimens collected in Kobe Bay ( Japan), later reported from the same area by Imajima and Hartman (1964) and described in more detail by Imajima and Williams (1985). Surprisingly, this work ignores the record of Williams (1984) in the Indian Ocean ( Mozambique Channel) probably due to simultaneous date of publication. Our specimens clearly differ from T. kobei in having notopodia and notochaetae of CH 1 much longer (similar to Williams’ material), by the absence of a conspicuous triangular lobe and a glandular area at the level of notopodia of CH 3 (Table 1) and by the similar length and high degree of fusion of posterior branchial lobes (see also Hutchings & Peart 2000, Table 3A).
Terebellides californica was described by Williams (1984) from the Pacific Ocean (Oregon to Western Mexico) at shelf and slope depths, and later reported from the same area by Hernández-Alcántara and Solís-Weiss (1999) and Hilbig (2000). Hernández-Alcántara and Solís-Weiss (1998) report T. californica for the Mexican Caribbean but we suspect that this may correspond to other species with large CH 1 as well. Williams (1984) characterized the taxon only by the “very well developed first chaetiger with greatly prolonged fine notosetae” (Williams 1984, p. 128). The species was later redescribed by Hilbig (2000) proposing a “trilobed structure of the peristomium” as potential new diagnostic character. This character was not observed in our specimens, but in our opinion its relevance should be taken with caution as its presence in the species has not been sufficiently assessed. Terebellides mediterranea spec. nov., differs from T. californica in branchial structure in addition to their very different geographical location—shelf and slope depths of the Pacific Ocean in T. californica vs. shallow waters in the Adriatic Sea in T. mediterranea spec. nov. In this sense, in T. mediterranea spec. nov., a large fifth branchial lobe is present and posterior pairs of lobes (first to fourth lobes) are fused along most of their lengths while T. californica lacks a fifth branchial lobe and posterior pairs of lobes are moderately fused (Hutchings & Peart 2000; Schüller & Hutchings 2010).
Habitat. Offshore stations in the northern Adriatic Sea on silty sand bottom on 31 m depth. Known only from type locality.
Distribution. Adriatic Sea.
Etymology. The name of the species refers to the Mediterranean Sea.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |