
Amaralia oviraptor, Friel, John P. & Carvalho, Tiago P., 2016
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4088.4.4 |
|
publication LSID |
lsid:zoobank.org:pub:B116A7AF-58B1-4C86-A429-B24D7FACC431 |
|
DOI |
https://doi.org/10.5281/zenodo.5687306 |
|
persistent identifier |
https://treatment.plazi.org/id/038B0845-2D5B-E14A-76E0-FF28FA56EEFF |
|
treatment provided by |
Plazi |
|
scientific name |
Amaralia oviraptor |
| status |
sp. nov. |
Amaralia oviraptor , new species
( Figs. 1 View FIGURE 1 , 2 View FIGURE 2 , 3 View FIGURE 3 a, 4, 5, 6, 7a; Table 1 View TABLE 1 )
Amaralia hypsiura non (Kner, 1855). Castelo et al., 1978: 131 [illustrated and briefly described]. Britski et al., 1999: 111 [illustrated and briefly described]. Chernoff & Willink, 2000: 86 [listed]. Liotta et al., 2001: 4 [illustrated and briefly described]. Ferriz & Gómez, 2002: 1 [briefly described].]. Friel, 2003: 261 [referred as undescribed]. Veríssimo et al., 2005: 5 [listed]. Langeani et al., 2007: 13 [referred as undescribed]. Britski et al., 2007: 137 [illustrated and briefly described]. Polaz et al., 2014: 127 [listed].
Amaralia hypsiurus non (Kner, 1855). López et al., 2003: 63 [listed].
Amaralia sp. da Graça and Pavanelli, 2007: 109 [illustrated and briefly described]. Almirón et al., 2008: 139 [illustrated and briefly described]. Arias et al., 2013:14 [listed].
Bunocephalus hypsiurus non Kner, 1855. Mees, 1988: 91 [listed in part].
Holotype. MZUSP 4423, 68 mm SL, Brazil, Mato Grosso, Santo Antônio do Leverger municipality, rio Cuiabá, 15°52'S 56°05'W, 1965, G. Olson.
Paratypes. ANSP 197190, 3 (1 c&s), 104–119 mm SL, Paraguay, Cordillera, Río Piribebuy near confluence with Río Paraguay ca. 4 mi northwest of Emboscada, 25°04'09''S 57°23'03''W, Feb 2014, Agustin Villanucci. LBP 12968, 1, 107 mm SL, Brazil, Mato Grosso, Poconé municipality, rio Cuiabá at Parque Nacional do Pantanal Matogrossense, 17°49'51''S 57°23'42''W, 25 Oct 2010, R. Britzke, L. R. Gaspar & B. F. Melo. MNRJ 20329, 1, 31.4 mm SL, Brazil, Mato Grosso, Pedra Preta municipality, rio Jurigue on highway BR-364 near Pedra Preta, tributary to rio Vermelho, 16°37'26''S 54°26'22''W, 13 Feb 2000, Melo, P. A. Buckup & Melo. MNRJ 27700, 103 mm SL, Brazil, Mato Grosso, Porto Esperidião municipality, baía do Campo at fazenda Pantanalzinho, 15°51'S 58°27'W, 27 Nov 1984, D. F. Moraes Jr. and G. W. A. Nunan. MZUSP 36383, 1, 100.7 mm SL, Brazil, Mato Grosso do Sul, Corumbá municipality, 19°00'S 57°39'W, Feb 1985, E. K. Bastos. MZUSP 41099, 1, 35.3 mm SL, Brazil, Mato Grosso do Sul, Corumbá municipality, rio Miranda, dead arm downstream Passo do Lontra, 20°14'S 56°22'W, 13 Sep 1989, E. K. de Resende. MZUSP 49024, 2, 66–88 mm SL, Brazil, Mato Grosso do Sul, Corumbá municipality, rio Miranda at Base de Estudos do Pantanal, 20°14'S 56°22'W, 25 Jan 1992, O. Froehlich. MZUSP 59635, 1, 32 mm SL, Brazil, Mato Grosso do Sul, Rio Negro municipality, rio do Peixe on highway MS-080, 19°23'25''S 54°59'19''W, 26 Aug 1998, N. Menezes & B. Chernoff. NRM 15916, 1, 82.7 mm SL, Paraguay, Amambay, Arroyo Aquidaban-Nigui at monument site and slightly upstream in Parque Nacional Cerro Corá, 22°38'S 56°01'W, 25 Jan 1992, S. O. Kullander et al. NRM 15917, 1, 59.5 mm SL, Paraguay, San Pedro, Río Aguaray-Guazú at Lima, 23 Jan 1992, S. O. Kullander et al. NRM 33073, 2, 58.8–119 mm SL, Paraguay, Misiones, Villa Florida, Río Tebicuary at Centu-Cué, 26°24'42''S 57°02'38''W, 4 Dec 1995, S. O. Kullander et al. UMMZ 206795, 1, 46.8 mm SL, Paraguay, Amambay, Río Apa ca. 0.5 km upstream of bridge between Brazil and Paraguay at Bella Vista, 22°06'S 56°31'W, 27 Jul 1979, J. N. Taylor, T. Grimshaw, R. Myers et al. UMMZ 207818, 3 (1cs), 84.3–118 mm SL, Paraguay, Concepción, Río Aquidaban at Paso Horqueta ca. 24 km NNW of Loreto, 23°03'S 57°23'W, 5 Sep 1979, J. N. Taylor, G. Smith, B. Smith, E. Koon, and R. Myers. UMMZ 216543, 1, 66.5 mm SL, Paraguay, Central, Río Paraguay at Playa Carrasco-Guaradero ca. 5 km N of port of Asunción, 25°15'S 57°37'W, 24 Aug 1980, L. Naylor & O. Romero.
Sequences. Genseq-2 COI: We published sequences of the mitochondrial gene cytochrome oxidase subunit one (COI; Hebert et al., 2003) of 2 paratypes of Amaralia oviraptor , for which tissue samples fixed in 96% alcohol were available. These sequences correspond to genseq-2 COI following the nomenclature of Chakrabarty et al. (2013). GenBank accession numbers for these specimens are: KT820070 View Materials and KT820071 View Materials for LBP 12968 and ANSP 197190, respectively. In addition, we sequenced the same mitochondrial gene for two specimens of Amaralia hypsiura from the Xingu River drainage ( KT820068 View Materials for ANSP 197607 and KT820069 View Materials for INPA 47820).
Diagnosis. Amaralia oviraptor is distinguished from its only congener, Amaralia hypsiura , by the higher number of dorsal-fin rays (3 vs. 2; Fig. 3 View FIGURE 3 ); by the presence of a lateral contact between middle and posterior nuchal plates (vs. middle and posterior nuchal plates not contacting each other laterally, Fig. 3 View FIGURE 3 ); by the presence of feeble serrations along the distal portion of the anterior margin of pectoral-fin spine ( Fig. 7 View FIGURE 7 A vs. absence of serrations along the anterior margin; Fig. 7 View FIGURE 7 B); and by a longer cleithral process (17.4–19.5 % of SL, mean 18.2 % vs. 14.0– 17.2 % of SL, mean=15.5 %).
Description. Morphometric data summarized in Table 1 View TABLE 1 . Maximum body size moderate to large compared to other aspredinids (maximum observed size 1228 mm SL). Dorsal, left lateral and ventral views of holotype shown in Figure 1 View FIGURE 1 . Anterodorsal view of a live paratype specimen shown in Figure 2 View FIGURE 2 . Head and anterior body depressed, lateral profile ascending from tip of snout to vertical with pelvic-fin origin, with convex bony prominences in between. Posterodorsal profile of body ascending at vertical with pelvic-fin origin and somewhat straight after this point to base of caudal fin, but bearing convex knobs on dorsal portions of neural spines. Ventral body profile convex from mouth to branchial aperture, almost straight from this point to base of caudal fin, but bearing convex bony knobs on ventral portion of caudal peduncle. Convex from pectoral girdle to anus in specimens with filled and dilated stomachs. Caudal peduncle deep, and compressed laterally.
Skull ornamentation strongly developed, pronounced bony knobs on anterolateral portion of mesethmoid, antorbital, frontal posterior to eye, lateral portion of sphenotic, anterocentral portion of pterotic and posterior tip of supraoccipital. Eye small and positioned dorsolaterally. Skin covering eye dense and pale. Anterior nostril located terminally at tip of snout, associated with fleshy tube projecting beyond upper lip. Posterior nostril without flap, opening anteriorly and near eye. Mouth subterminal, upper lip more prominent than lower lip. All barbels simple, unbranched; maxillary barbel just reaching branchial aperture; posterolateral mental barbel about twice as long as anteromedial mental barbel. Opercular opening reduced to small valvular slit located ventrally, anteromedially positioned to pectoral-spine insertion. Axial slit pore present, dorsoventrally inclined underneath posterior cleithral process. Adult males with digitiform testes. Integument covered with large unculiferous tubercles scattered throughout head and body, larger on dorsal and lateral portions. Tubercles forming series of larger and horizontally aligned rows on posterior portion of body with three well-defined rows on lateral of body, one at lateral line others just above and below.
Anterior margin of mesethmoid somewhat rounded, not anterolaterally projected, its dorsal surface strongly concave ( Fig. 4 View FIGURE 4 ). Contact between posterior margin of mesethmoid and frontal straight, not elevated. Ethmoid cartilage separate from articular facet of palatine. Frontal with anterolateral and posterolateral projections forming margin of orbit. Posterior projections of frontal absent, frontal contacting just sphenotic posteriorly and not supraoccipital. Frontal epiphyseal bar present, separating anterior and posterior cranial fontanels. Supratemporal fossa at contact between pterotic and supraoccipital bones. Pterotic with laterally expanded bony shelf, lateralmost portion rounded followed posteriorly by pronounced concavity. Premaxilla somewhat rectangular in shape, with teeth attached throughout most of its ventral surface. Dentary robust, deepest at coronoid process, tapering anteromedially ( Fig. 5 View FIGURE 5 ), abutting counterpart at medial portion. Dentary teeth arranged mostly in one row on its dorsal margin, up to three rows towards symphysis. Ascending process of Meckel’s cartilage hook-shaped, not contacting main portion of this cartilage. Coronomeckelian bone present. Hyomandibula associated with preopercle and posterior portion of preopercular-mandibular laterosensory canal, supraopercle absent. Cartilaginous articulation or joint of hyomandibula with neurocranium restricted to sphenotic bone. Anterodorsal process of hyomandibula developed, contacting ventral surface of sphenotic. Opercular condyle of hyomandibula well developed, directed posteroventrally. Metapterygoid present, contacting quadrate ventrally and variably contacting hyomadibula posteriorly. Endopterygoid present, somewhat rectangular in shape, located ventrally posterior to contact of palatine and lateral ethmoid. Posterior margin of autopalatine rounded in shape, presenting cartilage. Opercle “L” shaped, posterior arm longer than ventral arm. Interopercle present, somewhat triangular in shape and firmly attached to ventral arm of opercle. Dorsal hypohyal absent. Anterior ceratohyal with expanded blade on anteroventral margin, contacting posterior ceratohyal by means of cartilage within an interdigitated suture. Four branchiostegal rays. Urohyal with well developed lateral wings, without a medial foramen. First and second pharyngobranchials absent, third and fourth present and ossified. First hypobranchial ossified, second and third cartilaginous. Second and third basibranchial ossified, third cartilaginous. Third epibranchial bearing uncinate process. Gill rakers present on all ceratobranchials. Pharyngeal teeth well developed on upper tooth plate; a single row of teeth on medial margin of fifth ceratobranchial.
Dorsal lamina of Weberian apparatus reaching dorsal surface of body, lateral profile of lamina with concavity at about one third of its length. Enclosed aortic canal on the ventral surface of Weberian apparatus. Parapophysis of fourth vertebra forming broad lamina, contacting parapophysis of fifth vertebrae. Parapophysis of fifth vertebra long, extending to lateral body surface transverse to main body axis. Distal portion of fifth parapophysis not expanded. Vertebrae with horizontal transverse processes from centrum seven to 21–22 [19(1); 21(2); 22*(2)]. Vertebrae possessing opening for hemal canal at vertebra six and at vertebra 10. Hemal and neural spines on anterior portion of body vertically displaced relative to body axis, posteriorly inclined on caudal peduncle. Hemal and neural spine anteroposteriorly expanded and long, reaching dorsal surface of body. Hemal spines contacting anal-fin pterygiophores bifid. Total vertebrae 29–31 [29(2); 30*(2); 31(2)]. Four pairs of ribs, on vertebrae six to nine.
Dorsal fin with three rays, without spinelet ( Fig. 3 View FIGURE 3 A). First ray unbranched, spinous, followed by two branched rays. Last dorsal-fin ray free from back, not adnate. Anterior nuchal plate absent, middle nuchal plate contacting posterior nuchal plate laterally ( Fig. 3 View FIGURE 3 A). Posterior nuchal plate not developed laterally, lateral margin extending slightly beyond contact with middle nuchal plate. Pectoral fin with rigid spine and five to six (modally five) branched soft rays, except for last ray unbranched. Pectoral-fin spine slightly curved along its main axis, bearing fine ridges on ventral and dorsal surfaces of its shaft. Pectoral-fin tip bearing two to three short v-shaped unossified lepidotrichia, its hemitrichs of approximately same size ( Fig. 7 View FIGURE 7 ). Anterior margin of pectoral-fin spine with feeble serrae along its distal fourth, remaining portion of spine smooth. Posterior margin of spine bearing antrorse dentations on its distal half. Two ossified plus one cartilaginous pectoral-fin radial. Supracleithrum not fused with neurocranium, bearing large knobby ridge on its dorsal surface. Postcoracoid process moderate, extending slightly beyond the postcleithral process. Pelvic fin with six soft rays, third and fourth rays longest, not reaching anal-fin origin, first ray unbranched followed by four branched rays and last frequently unbranched. Basipterygium posterior margin jagged. Anal fin with five to six rays (modally five), first two or three unbranched, a third of length of last anal-fin ray extension adnate by membrane to body. Caudal fin with nine principal rays, five associated with upper lobe and four with ventral lobe (Fig. 6), posterior margin of caudal fin convex. Lowermost and uppermost caudal-fin rays unbranched, those slightly shorter than following branched rays. Caudal fin with two thickened procurrent rays on upper and lower lobes, anterior procurrent ray rod-shape or reduced and posterior procurrent ray thickened and “S” shape, sometimes partially fused with anterior procurrent ray. Posterior margin of upper hypural (hypurals three to five) extending posteriorly further than lower hypural plate (hypurals one and two fused with parhypural). Hypurapophysis developed at parhypural located ventrally to first preural centrum. Second ural half-centrum well developed (Fig. 6). Adipose fin absent.
FIGURE 6. Lateral view of left side of caudal skeleton of Amaralia oviraptor ANSP 197190, 104 mm SL. DPR, dorsal procurrent caudal-fin rays; FR, ventralmost unbranched caudal-fin ray; EP+UN, epural fused with uroneural; HS, hemal spine of 30th vertebra; HY3-5, hypurals 3–5; HYP, hypurapophyses; PH +HY1-2; parhypural fused with hypurals 1–2; PU1+U1, first preural centrum plus first ural centrum; U2, second ural centrum; VPR, ventral procurrent caudal-fin rays. Scale bar = 2mm.
Nasal bone ossified, lateral to mesethmoid. Infraorbital 1 anterior limb pointed, extending anterior to anterior margin of premaxilla. Antorbital mesial limb rounded and associated with infraorbital lateralis canal. Three additional tubular infraorbital ossifications, passing below eye margin and entering neurocranium through sphenotic. Mandibular canal incomplete and not enclosed in dentary bone. Mandibular canal bearing a single tubular ossification at vertical with retroarticular bone, a gap, and posteriorly three to four tubular ossifications near its entrance in preopercle ( Fig. 5 View FIGURE 5 ). Extrascapular present ( Fig. 4 View FIGURE 4 ). Anterior portion of lateral line running just aside lateral margin of fourth parapophysis. Lateral line complete, straight and extending to hypural plate, formed by simple tubes with lateral hook-shaped projections and without dorsal and ventral plate-like extensions.
Color in alcohol. Head and body light brown to dark gray, venter of body about as dark as dorsum ( Fig. 1 View FIGURE 1 ). A series of light spots on distal portions of enlarged tubercles scattered over the body, these most conspicuous and enlarged in the series of tubercles associated with the lateral line. All fins mostly dark with clear distal portions, proximal portions of pelvic fin with light scattered areas. Live specimens have similar color pattern ( Fig. 2 View FIGURE 2 ).
Distribution. Amaralia oviraptor is widely distributed in the Paraná-Paraguay River system, found throughout the Paraguay River Basin in Brazil, Paraguay and northern Argentina and also in the Paraná drainage in Argentina and Brazil ( Fig. 8 View FIGURE 8 ). The new species seems to be absent in some portions of this system such as the Uruguay River Basin and the main tributaries of the upper Paraná.
Etymology. The epithet oviraptor is a combination of the Latin ovum (ovi), meaning egg; and raptor, a robber or plunderer, commonly used term for a predator, here referring to the peculiar dietary preferences observed in this species. Treated as a noun in apposition.
Ecological notes. An interesting aspect of Amaralia is its apparent dietary specialization on both the eggs and developing embryos of other catfishes (Friel, 1994; Roberts, 2015). Direct evidence of such oophagy is based on examination of the stomach contents from 23 preserved Amaralia specimens (17 A. hypsiura and six A. oviraptor ). Seven of these specimens (six female and one male) contained masses of ova or developing embryos in their stomach. The source of the eggs and embryos is most likely those of loricariid catfishes, and in least in one instance, the caudal fin-ray counts of the embryos fall within the range for loricariid catfishes. One male contained a single scoloplacid catfish specimen in its stomach, and the remaining 15 specimens (seven female and eight male) had empty stomachs. These observations differ significantly from the documented diets of other aspredinids, where stomach contents typically contained some organic detritus along with various aquatic and terrestrial insect prey.


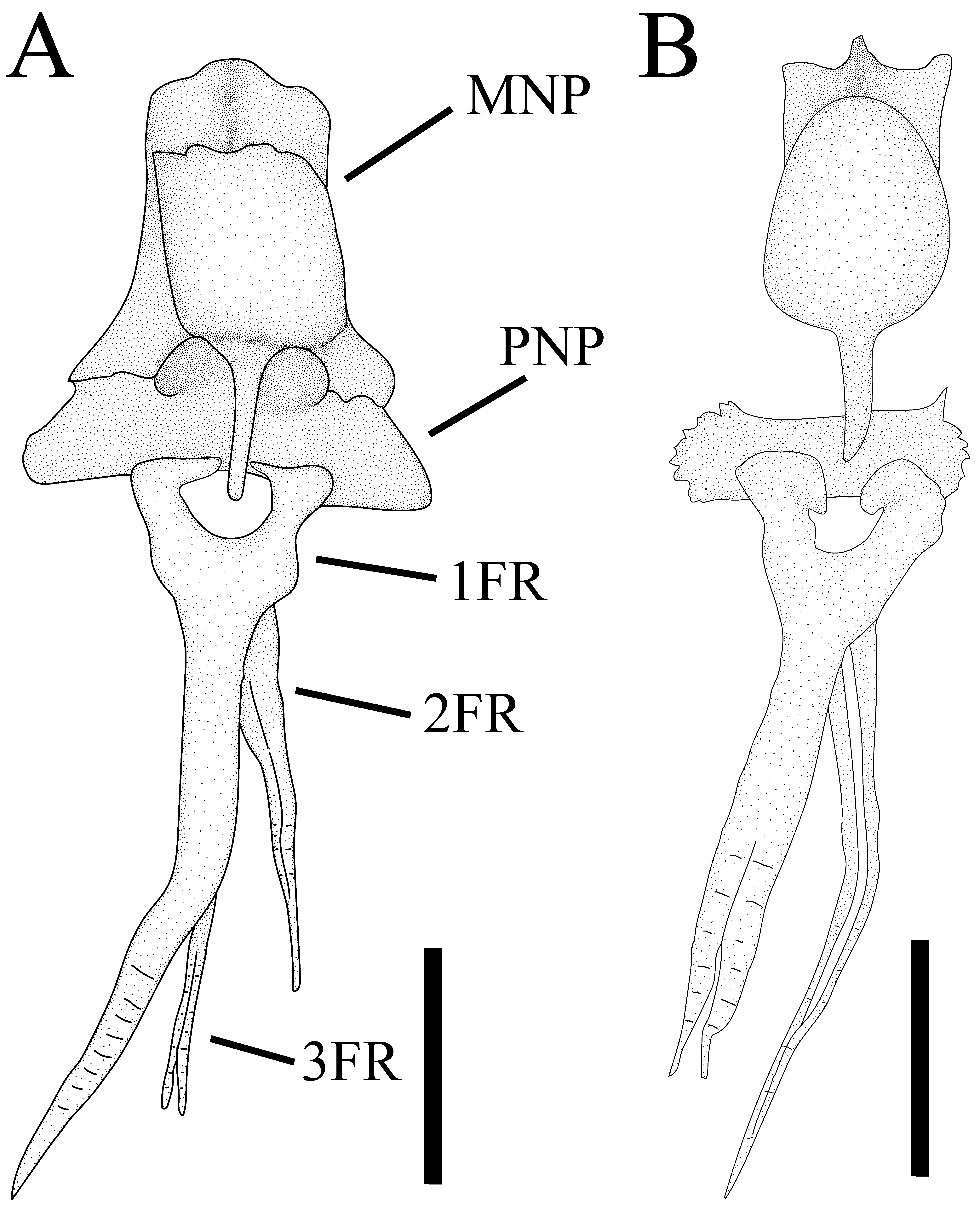

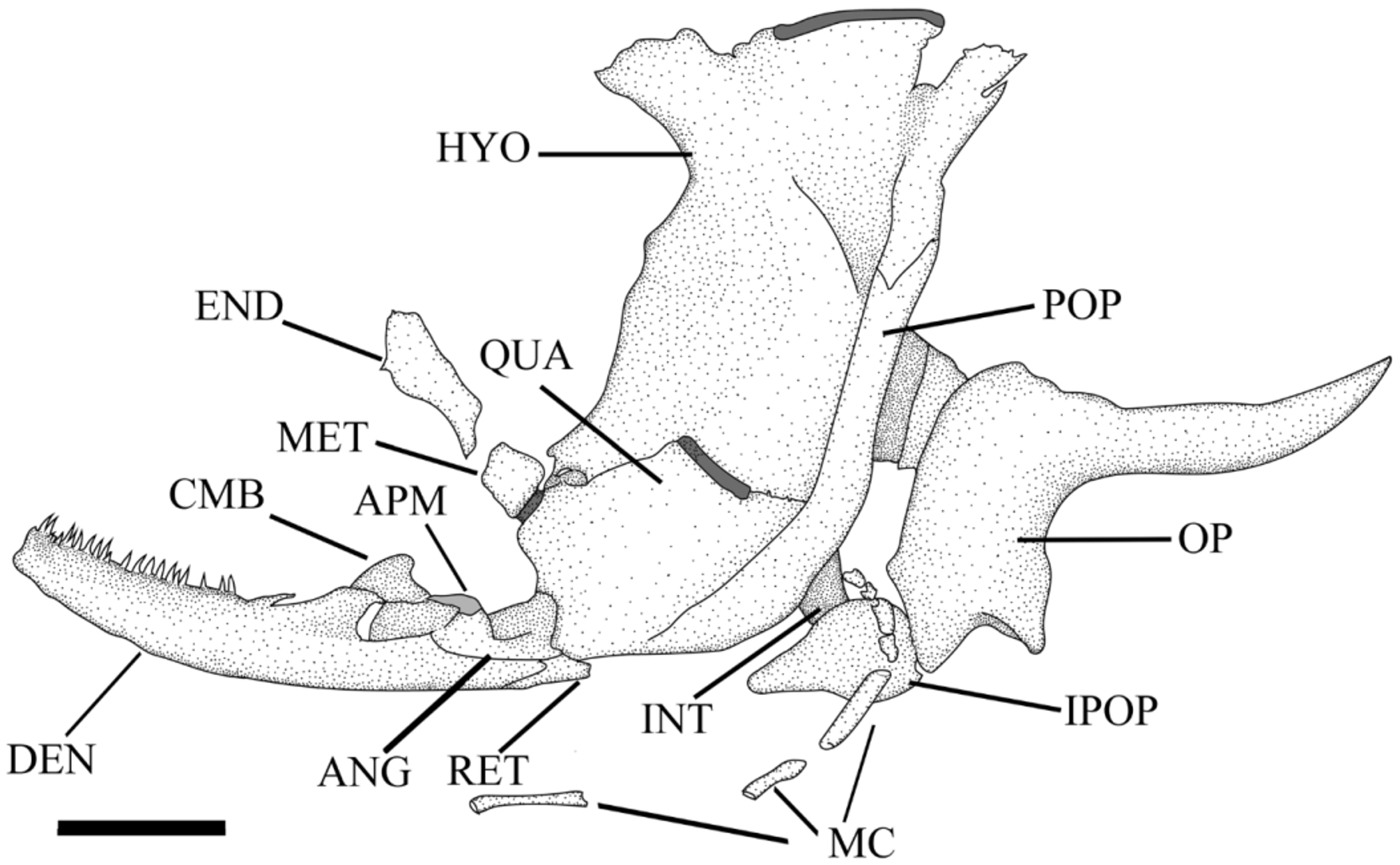


TABLE 1. Morphometric data of holotype (H) and paratypes of Amaralia oviraptor (n = 12 including the holotype) and Amaralia hypsiura (n = 14). SD = standard deviation.
| Amaralia oviraptor | Amaralia hypsiura | ||||
|---|---|---|---|---|---|
| H | Min Max Mean | SD | Min Max Mean | SD | |
| Standard length | 68.4 | 35.6 122.8 78.13 | - | 34.7 132.0 76.46 | - |
| Percent of standard length | |||||
| Head length | 23.5 | 22.0 27.6 23.6 | 1.67 | 21.6 25.4 23.0 | 1.07 |
| Prepectoral length | 25.3 | 24.1 30.1 26.0 | 1.67 | 23.7 29.9 26.4 | 1.57 |
| Cleithral width | 32.4 | 30.9 34.6 32.8 | 1.11 | 29.5 36.9 33.2 | 2.41 |
| Maximum head depth | 12.6 | 11.0 15.7 12.8 | 1.26 | 10.3 18.1 13.9 | 2.30 |
| Pectoral-spine length | 26.6 | 21.3 27.2 25.0 | 1.93 | 17.9 27.9 22.9 | 3.12 |
| Distance between coracoid processes | 21.1 | 18.8 25.3 20.8 | 1.90 | 19.9 26.8 22.7 | 2.33 |
| Coracoid process length | 13.9 | 12.5 15.4 13.6 | 0.86 | 9.7 16.0 12.5 | 1.88 |
| Distance between cleithral processes | 25.0 | 22.6 27.8 25.4 | 1.47 | 24.3 33.1 28.0 | 2.62 |
| Cleithral process length | 17.9 | 17.4 19.5 18.2 | 0.64 | 14.0 17.2 15.5 | 0.92 |
| Predorsal length | 44.9 | 41.1 44.9 42.9 | 1.17 | 39.6 47.6 41.9 | 2.23 |
| Depth at dorsal-spine insertion | 12.4 | 10.0 14.8 12.7 | 1.24 | 10.6 20.7 15.0 | 2.90 |
| Length of unb 1 st dorsal-fin ray | 8.6 | 5.3 9.1 7.9 | 1.05 | 5.0 18.9 14.8 | 4.10 |
| Prepelvic length | 52.2 | 49.8 55.4 52.8 | 1.54 | 51.8 57.7 53.7 | 1.80 |
| Length of unb 1 st pelvic-fin ray | 11.7 | 9.9 13.2 11.7 | 1.13 | 9.6 13.2 11.5 | 1.18 |
| Preanal length | 68.6 | 64.7 73.1 68.9 | 2.29 | 66.8 73.6 69.4 | 1.76 |
| Anal-fin base length | 13.6 | 10.1 15.5 13.9 | 1.43 | 11.3 15.5 13.2 | 1.22 |
| Caudal-peduncle length | 17.1 | 16.6 22.5 18.8 | 1.94 | 15.0 20.9 18.3 | 1.55 |
| Caudal-peduncle depth | 10.9 | 9.5 11.0 10.3 | 0.53 | 10.2 15.1 12.7 | 1.51 |
| Caudal length | 20.6 | 18.0 23.3 21.1 | 1.65 | 17.3 21.5 19.0 | 1.67 |
| Percent of head length | |||||
| Snout length | 26.7 | 23.1 30.6 26.7 | 2.01 | 22.9 31.1 28.1 | 2.28 |
| Eye diameter | 8.7 | 7.7 12.9 9.5 | 1.57 | 6.7 14.1 9.7 | 2.08 |
| Interorbital width | 27.3 | 25.5 34.7 28.5 | 2.42 | 20.4 37.6 28.9 | 4.71 |
| Maxillary-barbel length | 78.9 | 55.2 80.3 72.1 | 8.39 | 46.8 76.9 62.6 | 9.06 |
| Distance between anterior nares | 13.7 | 12.8 19.0 16.3 | 1.78 | 11.5 23.3 16.6 | 2.79 |
| Distance between posterior nares | 34.8 | 30.4 40.1 35.0 | 2.43 | 27.4 43.3 35.4 | 3.81 |
| Mouth width | 33.6 | 28.0 43.5 37.4 | 3.98 | 24.2 41.8 35.9 | 5.00 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |