
Fagaroxylon undetermined
|
publication ID |
https://doi.org/ 10.5070/P9401462457 |
|
persistent identifier |
https://treatment.plazi.org/id/038AF505-A318-9608-553C-FA5CFA549F00 |
|
treatment provided by |
Felipe |
|
scientific name |
Fagaroxylon undetermined |
| status |
|
CF. FAGAROXYLON SP.
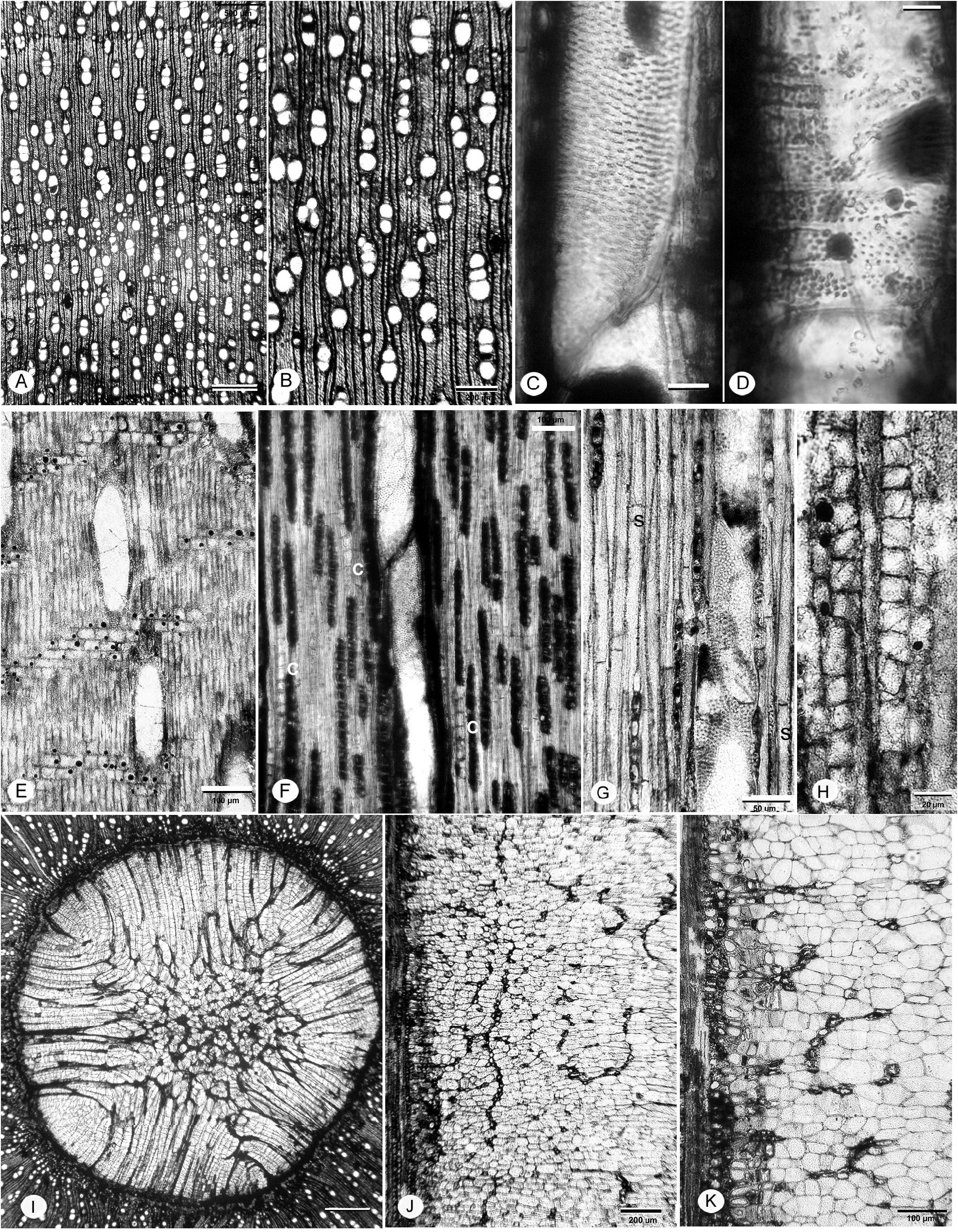
FIG. 18A–I View Figure 18
Description— Growth rings present, marked by radially narrow fibers, (probably also marginal parenchyma) ( Fig 18A, B View Figure 18 ).
Wood diffuse-porous ( Fig 18A, B View Figure 18 ). Vessels solitary (14%) and in radial multiples mostly 2–4 (-7), clusters rare; mean tangential diameter 63 ( SD =15) µm, range 28–90 µm; 36–42–50 vessels/mm 2. Perforation plates simple ( Fig. 18E, F View Figure 18 ); intervessel pits crowded alternate ( Fig. 18C View Figure 18 ), horizontal diameters small to medium 5–8.5 µm. Vessel-ray parenchyma similar to intervessel pits ( Fig. 18D View Figure 18 ). Vessel element lengths 235–277–353 µm (n=12); vessel element end walls obliquely inclined. Helical sculpturing (grooves) in some narrow vessel elements ( Fig. 18E, F View Figure 18 ). Gum-like deposits common.
Fibers thin-walled, non-septate, pitting not observed ( Fig. 18E–H View Figure 18 ).
Axial parenchyma not common, scanty paratracheal (hard to distinguish from thin-walled fibers) and prob- ably also marginal ( Fig. 18H View Figure 18 ).
Rays 1–4 (-5)-seriate ( Fig. 17E, G View Figure 17 ). Average ray height 497 ( SD =203) µm, range 181–1062 µm. Homocellular, composed of procumbent cells ( Fig. 18D, F, I View Figure 18 ), 6–9 per mm, non-storied.
Crystals common in chambered axial cells (fibers), strands sometimes adjacent to rays, one crystal per chamber, numerous (10+) crystalliferous chambers per strand ( Fig. 18E, G, I View Figure 18 ).
Specimen— UF 278-84907, estimated maximum diameter 10 cm.
Occurrence— Dietz Hill ( UF 278).
Comparisons with extant woods— We searched the InsideWood database multiple times. One search used diffuse porosity (5p), absence of vessel patterning and exclusively solitary vessels (6-9a), exclusively simple perforation plates (13p 14a), alternate intervessel pitting that is neither minute nor large (22p, 24a, 27a), vessel-ray parenchyma pits similar to intervessel pits (30p), average vessel diameter less than 200 µm (43a), vessels per mm 2 between five and 100 (46a, 50a), non-septate fibers with simple to minutely bordered pits (61p 66p), axial parenchyma scanty paratracheal and marginal, but not accompanied by more obvious axial parenchyma (78p, 89p, 80-88a), homocellular rays not exclusively uniseriate or greater than 10-seriate (96a, 99a, 104p), storied structure absent (118–122a), oil/mucilage cells and intercellular canals absent (124–130a), prismatic crystals present in fibers (136p 143p). When not allow- ing any mismatches, this search yielded only Meliaceae ( Capuronianthus J.- F.Leroy, 1958; Quivisianthe Baill., 1893 , from Madagascar ( Rutaceae ); Aeglopsis Swingle, 1912 ; Afraegle (Swingle) Engl., 1915 , from Africa; and Sapindaceae (one Asian Acer L., 1753).
When one mismatch was allowed, results again included Meliaceae (three species), Rutaceae (12 species), Sapindaceae (19 species, including 11 of Acer ). Other families retrieved were Combretaceae R.Br.(1810) (three Terminalia L. (1767) species), which differ in having crystals in idioblasts or in having more abundant axial parenchyma), Lamiaceae (one Vitex species with mostly septate fibers and crystals of other shapes), Fabaceae (five species, none have crystals in fibers, although there is the possibility crystals in fibers might have been interpreted as crystals in chambered axial parenchyma as reported for four of them).
We tried other searches using slightly different combinations of features, and all of them consistently returned members of the Sapindalean families Meliaceae , Rutaceae , and Sapindaceae . We are confident that this wood belongs to the Sapindales and are inclined to think that it is most likely related to Sapindaceae , which is noted as the Sapindalean family most commonly having crystals in fibers ( Pace et al. 2022). It resembles Acer most closely, but all present-day Acer consistently have helical thickenings in vessel elements, so this wood is not assigned to Acer . The fruit and seed flora of Dietz Hill has an endocarp that might have affinities with Acer ( Manchester and McIntosh 2007).
Dipteronia Oliv. (1889) is closely related to Acer and fruits of it occur in the Teater Road shales that we infer to be of similar age to the Dietz Hill ( UF 278) local- ity ( McClain and Manchester 2001). Itoh et al. (2022) described Dipteronia sinensis Oliv. (1889) (under the name Acer dielsii H. Lév. (1912) . That description differs somewhat from the descriptions and observations of D. dyeriana A. Henry (1903) and D. sinensis made by Peter Gasson (Royal Botanic Gardens, Kew) and Michael Wiemann (Center for Wood Anatomy Research, Madison) and available on InsideWood (insidewood.lib.ncsu.edu). Itoh et al. (2022) report helical thickenings in narrow vessel elements and rays commonly more than 1 mm high, features not seen by Gasson or Wiemann. Crystals are not reported for Dipteronia .
Comparisons with fossil woods— Excluding fossil wood descriptions with most features coded as unknown, the results of our searches of InsideWood’s fossil wood database returned two species of Fagaroxylon Van der Burgh (1964) from the Miocene of Europe. Fagaroxylon is a genus created for woods with features of both Fagara L. (1759), which is now subsumed into Zanthoxylum L. (1753). Its diagnosis ( Van der Burgh 1964) and later descriptions of Fagaroxylon bavaricum ( Selmeier 1975) and Fa. limburgense ( Van der Burgh 1973) are remarkably similar to this Dietz Hill ( UF 278) wood. Similarities include most vessel features, axial parenchyma distribution, chambered crystals, ray width and cellular composition, and even the occasional occurrence of helical vessel wall thickenings. Vessel frequency is lower in Fa. limburgense , but this likely is because in the past vessel multiples (groups) were counted as one unit ( Wheeler 1986). Fagaroxylon limburgense , the type species, is reported to have fibers with distinctly bordered pits (diameter 3 µm) in their radial walls, a feature we did not observe in UF 278-84907. Because variations in preservation and the effects thereof can make it difficult to determine whether fiber pits are>3 µm across, we choose to assign UF 278-84907 to Fagaroxylon .
Fagaroxylon ’s combination of features occurs in other genera of Rutaceae , as well as the Meliaceae and Sapindaceae and is not unique to Zanthoxylum . It should be noted that seeds that correspond closely to the extant genus Zanthoxylum occur at Dietz Hill ( UF 278).
Other Sapindalean fruit and seed remains in the Teater Road shales include Koelreuteria Laxm (1771) ( Wang et al 2013), Dipteronia ( McClain and Manchester 2001), and several species of Acer leaves and fruits ( Wolfe and Tanai, 1987, indicated as “Sheep Rock Creek”).

| SD |
San Diego Natural History Museum |
| UF |
Florida Museum of Natural History- Zoology, Paleontology and Paleobotany |
| J |
University of the Witwatersrand |
| F |
Field Museum of Natural History, Botany Department |
| L |
Nationaal Herbarium Nederland, Leiden University branch |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |