
Plerandreoxylon oskolskii, Wheeler & Manchester & Baas, 2023
|
publication ID |
https://doi.org/ 10.5070/P9401462457 |
|
persistent identifier |
https://treatment.plazi.org/id/038AF505-A316-960D-57D7-F8A2FAD69EE3 |
|
treatment provided by |
Felipe |
|
scientific name |
Plerandreoxylon oskolskii |
| status |
sp. nov. |
PLERANDREOXYLON OSKOLSKII SP. NOV.
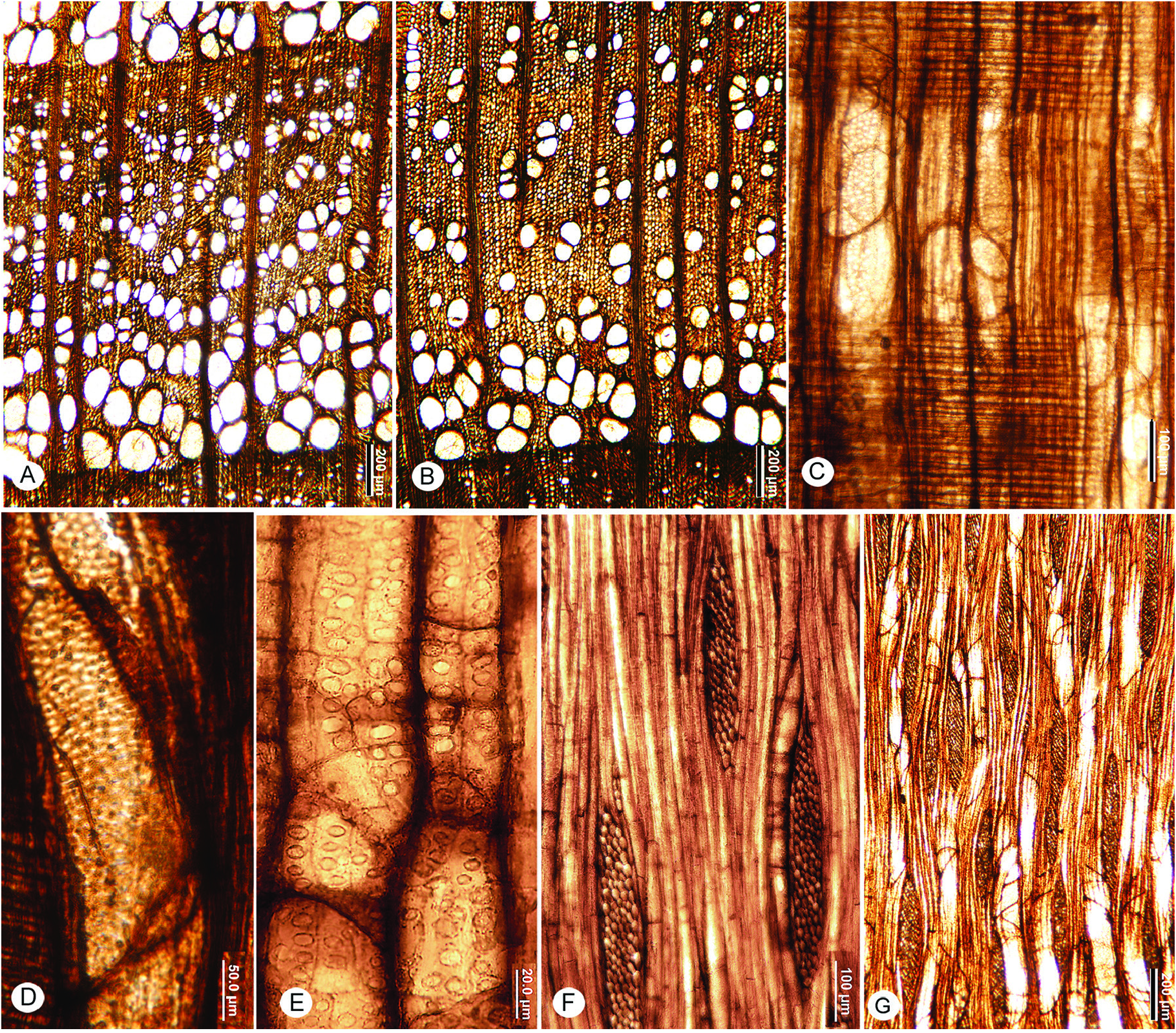
FIG. 19A–G View Figure 19
Diagnosis— Growth rings distinct, marked by radially flattened latewood fibers, and differences in vessel diameter between latewood and earlywood of subsequent rings.
Semi-ring-porous to ring-porous. Vessels solitary and in radial multiples, small clusters in the latewood. Earlywood with multiple rows of vessels, latewood vessels arranged in a tangential to diagonal pattern in narrower rings. Perforation plates simple, intervessel pits alternate, medium to large; vessel-ray parenchyma pits with reduced borders to simple, oval to slightly horizontally elongate. Tyloses present.
Fibers with simple to minutely bordered pits; thin to thick-walled; septate.
Axial parenchyma extremely rare.
Rays predominantly multiseriate, sheath cells occasional, average ray height <1 mm. Rays homocellular, composed of procumbent body cells, or heterocellular with a single marginal row of square cells and weakly procumbent cells. Rays per mm few.
Holotype — UF 278-84872 , estimated maximum diameter 8–9 cm.
Paratype — UF 278-84905 .
Occurrence— Dietz Hill, UF 278.
Etymology— Named for our good colleague and friend Alexei Oskolski who has studied Araliaceae wood and done much to further the field of wood anatomy in general.
Description— Growth rings distinct, marked by radially flattened latewood fibers. Ring-porous to semi-ring-porous. Vessels solitary and in short radial multiples, small clusters in the latewood. Earlywood with multiple rows of vessels; latewood vessels arranged in a tangential to diagonal pattern in narrower rings, mostly in radial multiples, solitary vessels not common ( Fig. 19A, B View Figure 19 .). Earlywood vessels 97 ( SD =22) µm, range 48–137 µm. Vessel element lengths commonly 350–500 µm. Perforation plates simple ( Fig. 19C View Figure 19 ). Intervessel pits alternate, 8–10 µm in horizontal diameter, tending to be angular in outline ( Fig. 19D View Figure 19 ); vessel-ray parenchyma pits with reduced borders and oval to slightly horizontally elongate ( Fig. 19E View Figure 19 ). Tyloses present ( Fig. 19C, E, G View Figure 19 ).
Fibers with simple to minutely bordered pits, septate ( Fig. 19F View Figure 19 ), thin to thick-walled.
Axial parenchyma extremely rare.
Rays homocellular, composed of procumbent body cells, or heterocellular with a single marginal row of square cells and barely procumbent cells; sheath cells occasional; 3–7 cells wide, mostly 5–6 cells ( Fig. 19F, G View Figure 19 ). Ray height averages 449 (126) µm, range 126–728 µm; 3–4 rays per mm.
Crystals, oil/mucilage cells, storied structure, and radial canals absent.
Comparisons to extant and fossil woods— We searched the InsideWood database multiple times. One search used: distinct growth ring boundaries (1p); wood ring-porous to semi-ring porous (5a, not diffuse-porous); vessels in wavy tangential bands (6p);simple perforation plates (13p); intervessel pits alternate and not minute (22p 24a); vessel-ray parenchyma pits rounded with much reduced borders (30a, 31p); septate fibers (65p); axial parenchyma rare (75p); rays more than 4-seriate, <1 mm high and with one row of square marginal cells (98p 102a 106p); storied structure, oil/mucilage cells, and radial canals absent (118a 120a 124a 125a 126a 130a). The IW matches only included Araliaceae (order Apiales ): two modern ( Aralia elata var. ryukyuensis J.Wen (1994) and Kalopanax septemlobus (Thunb.) Koidz. (1925) , both East Asian) and two fossil woods ( Plerandreoxylon oregonensis Wheeler and Manchester, 2002 , and Aralinium excellens Platen, 1908 ), both from the Eocene of the USA. These two Dietz Hill woods' ab- sence of uniseriate rays and the occasional occurrence of sheath cells are additional features that are consistent with the Araliaceae . Unfortunately, Platen’s Aralinium excellens cannot be placed with certainty in the Araliaceae and reasonably compared with this Dietz Hill ( UF 278) wood because its description lacks details on ray cellular composition and height and whether fibers are septate or non-septate ( Platen 1908). Moreover, it was described as having crystals in enlarged cells, a feature not reported for the Araliaceae .
Another search used absence of vessel-ray parenchyma pits similar to intervessel pits (30a) rather than 31p and absence of obvious axial parenchyma features (77a, 79–86a) rather than 75p. This search returned an additional two modern woods ( Aralia spinosa L., 1753; Gamblea ciliata C.B. Clarke, 1879 ; Araliaceae ) and an additional fossil wood ( Evodia gadjirian Gregory, 1976 , from an unspecified Eocene site in the vicinity of Post, Oregon, USA). Unfortunately, the description of the putative Evodia Forst. and Forst. (1775) lacked information on intervessel and vessel-ray parenchyma pitting and ray cellular composition; the accompanying images were at too low of a magnification to determine those details. Whether or not this wood truly is Evodia ( Rutaceae ) or might be Araliaceae is impossible to determine because the location of Gregory’s material is unknown and so it cannot be reexamined.
Alternative searches, e.g., 1p, 4p, 9a, 10a, 13p, 26p, 30a, 31p, 56p, 61p, 65p, 98p, 106p, in addition to Araliaceae , also retrieved two extant species of Toricellia DC (1830) ( Toricelliaceae Hu 1934 ), also in the Apiales . Adams (1949, p. 239) remarked “[the] overall character of the wood is suggestive of araliaceaeous affinities.” Toricellia differs from the fossil in having helical vessel wall thickenings, taller rays (to 1.8 mm), slightly more rays per mm, scanty paratracheal parenchyma more obvious, and conspicuous tyloses ( Noshiro and Baas 1998). Toricellia DC , is known by fruits from the Clarno Nut Beds ( Manchester et al. 2017) but is not known from any Post locality.
There are araliaceous fruits ( Paleopanax oregonensis Manchester, 1994) and two species of fossil wood, Plerandreoxylon nutbedensis Wheeler and Manchester (2002) and Pl. oregonensis Wheeler and Manchester (2002) , known from the middle Eocene Clarno Formation. This Dietz Hill wood agrees with the diagnosis of Plerandreoxylon , aside from “rays usually less than 4-seriate” and so we have emended the diagnosis to rays less than 10-seriate. Plerandreoxylon nutbedensis differs in having some scalariform perforation plates, some scalariform intervessel pitting, with uniseriate rays more common; Pl. oregonensis has mostly non-septate fibers and some scalariform intervessel pitting ( Wheeler and Manchester 2002). We consider these differences sufficient to warrant assigning this wood to a new species of Plerandreoxylon .
? MALPIGHIALES JUSS. EX BERCHT. & J. PRESL, 1820
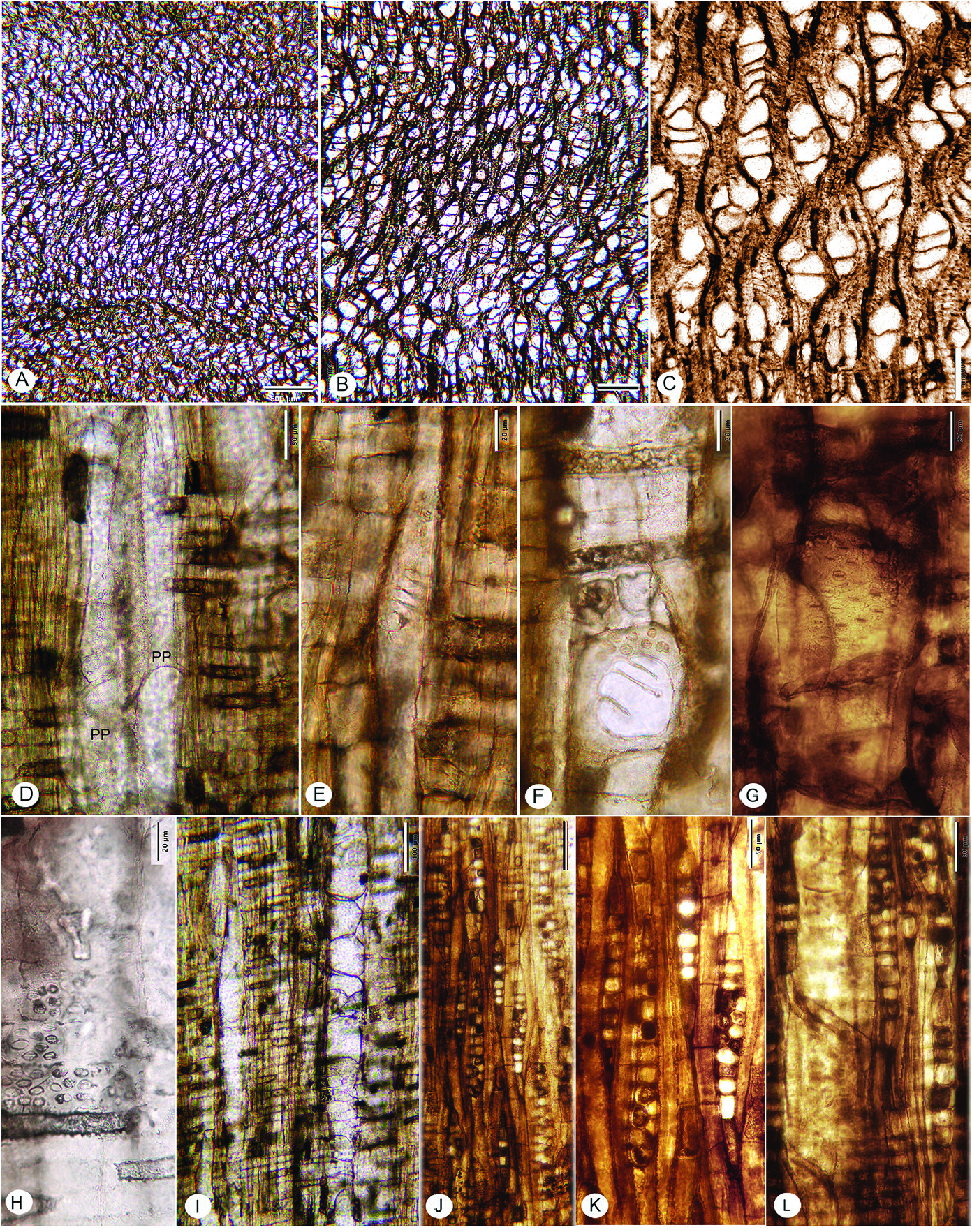
FIG. 20A–L View Figure 20
Description— Growth rings present, marked by radially narrow fibers ( Fig. 20A, B View Figure 20 ).
Wood diffuse-porous; solitary vessels rare, 8% solitary vessels, vessels predominantly in radial multiples of 2–6 (-9) ( Fig. 20A–C View Figure 20 ); mean tangential diameter 52 µm ( SD =11), range 26–76 µm; 90–121 vessels per mm 2. Perforation plates simple and scalariform with fewer than 10 bars ( Fig. 20D–F View Figure 20 ); intervessel pits alternate ( Fig. 20G View Figure 20 ), horizontal diameter 6–8–10 µm. Vessel-ray parenchyma pits similar in size to intervessel pits, generally oval in outline, with reduced borders ( Fig. 20H View Figure 20 ). Vessel elements short, <350 µm. Thin-walled tyloses abundant ( Fig. 20I View Figure 20 ).
Fibers thin- to thick-walled, apparently non-septate, pitting not observed.
Axial parenchyma not obvious in transverse sections, but visible in longitudinal section, strands of 4–8 cells.
Rays 1–2 seriate, mostly uniseriate ( Fig. 20J, K View Figure 20 ); average biseriate ray height 227 ( SD =52) µm, 131–315 µm. Rays composed primarily of procumbent cells ( Fig. 20I, L View Figure 20 ); 12–15 per linear mm.
Storied structure, radial canals, and oil/mucilage cells absent.
Specimen— UF 278-84892, estimated maximum diameter 10 cm.
Occurrence— Dietz Hill ( UF 278).
Comments— Specimen UF 278-84892 was problematic because it was highly compressed. The longitudinal sections provided a combination of radial and tangential views, measuring the quantitative vessel features and rays per mm were affected by the compression. The tyloses made it difficult to measure vessel element lengths.
Comparison with modern and fossil woods— We searched InsideWood’s modern and fossil wood database multiple times using various combinations of features, with almost all searches including wood diffuse-porous (5p), vessels in radial multiples of four or more common (10p), perforation plates simple (13p), intervessel pits alternate and not minute (22p 24a), fibers non-septate (66p), narrow rays (searching for rays 1–3 seriate - 97p or rays uniseriate - 96p). Although scalariform perforation plates are rare, we used their presence (14p). Most searches returned Nothofagaceae Kuprian (1962) ( Fagales ), which is an unlikely match because no macro- or microfossils of this Southern Hemisphere family have ever been found in the Northern Hemisphere ( Pujana et al. 2021). Members of the Salicaceae sensu APG were also returned as well as other malpighialean families (e.g., Achariaceae Harms, 1925 ; Euphorbiaceae Juss., 1789 ; Peraceae Klotzch, 1859 ; Phyllanthaceae Martinov, 1820 ; Putranjivaceae Endl., 1841 ). Results of other searches included Elaeocarpaceae Juss. (1816, Oxalidales Bercht. and J. Presl, 1820 ). No fossil wood in the InsideWood database was a good match. In spite of reviewing the family descriptions in Metcalfe and Chalk (1950) and reviewing the literature on the aforementioned families, at this time, we are not able to assign it to family. We suggest it belongs to the Malpighiales Martius , but we cannot confirm that placement.
Populus L. (1753) and Salix L. (1753) ( Salicaceae Mirbel (1815)) are common in the compression floras of the U.S.A., although woods of those genera are rare, which is probably because they readily decay making them less likely to enter the fossil record. Initially, we thought this sample might be Populus or Salix because the transverse section of this diffuse-porous wood shows narrow and abundant vessels that are commonly in radial multiples, abundant narrow rays, and axial parenchyma looked to be rare. However, this first impression was incorrect because this wood has some scalariform perforation plates, a feature that does not occur in present-day Salix or Populus . Moreover, strands of axial parenchyma are visible in the longitudinal sections. This sample serves as a cautionary tale that generally wood identification needs to be based on microscopic examination of longitudinal sections, not just transverse section.
| SP |
Instituto de Botânica |
| UF |
Florida Museum of Natural History- Zoology, Paleontology and Paleobotany |
| SD |
San Diego Natural History Museum |
| L |
Nationaal Herbarium Nederland, Leiden University branch |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |