
Halisarca dujardini
|
publication ID |
https://doi.org/ 10.5281/zenodo.206842 |
|
DOI |
https://doi.org/10.5281/zenodo.5626324 |
|
persistent identifier |
https://treatment.plazi.org/id/038A87FD-FFDD-FFD3-61C3-2626FCA1C4A4 |
|
treatment provided by |
Plazi |
|
scientific name |
Halisarca dujardini |
| status |
|
Redescription of Halisarca dujardini type species
( Figs. 2–23 View FIGURES 2 – 5 View FIGURE 6 View FIGURES 7 – 15 View FIGURES 16 – 19 View FIGURE 20 View FIGURES 21 – 23 )
Previous reviews. Lévi 1956; Bergquist 1996; Bergquist & Cook 2002.
Halisarca dujardini Johnston, 1842
Synonymy. Halisarca dujardini Johnston, 1842: 192 –193; Lendenfeld 1889: 729–730; de Laubenfels 1948: 175–176; Lévi 1956: 184; Chen 1976: 113–139; Ereskovsky 1993: 8; Bergquist 1996: 24–27; Bergquist & Cook 2002: 1090–1091. Halisarca franz-schulzei Merejkowsky 1878: 27–36, Pl. I, fig. 1–6; Pl. II, fig. 9–15. Hymeniacidon dujardini Bowerbank 1866: 224 . Halisarca sputum Topsent 1893: 38 –39.
Original description. Halisarca dujardini, Johnston, 1842 , p. 192.
Type material. Holotype: presumably lost. Neotype: BMNH 1960.1.7.1—Black Rock, Brighton, English Channel (epizootic on algae, collected close to the type locality) ( Bergquist & Cook 2002).
Comparative material examined. Sponge specimens used in the revision of Halisarca dujardini were collected using SCUBA, in White Sea (Kandalaksha Bay, Chupa Inlet (66°17'32"N – 33°39'39"E), 3–5 m depth, from algae Fucus ramosus and Laminaria saccharina , July 2000–2004; in Barents Sea, Kola Peninsula, Dalnije Zelentsi (69°07'19"N – 36°03'21"E) 0.3–0.5 m depth, July, August 2003; in Bering Sea, Avacha Bay (52°46'32"N – 158°36'16"E) 10-11 m depth, October 18 2008; in Japan Sea the Gulf of Peter the Great near of the Marine Biological station “Vostok” (Marine Biological Institute of Russian Academy of Sciences) (42°54'N – 132°38'W) from rocks and boulders at a depth 0.4–4 m; in North Sea, Norway, Bergen (60°23'29"N – 5°13'46"E), June 2009, 6 m depth; in North Sea, Netherlands, Zeeland, Goese Sas (51°38'52"N – 3°48'0"E) 18 m depth, July, 2009; NE Atlantic, North Wales (53°14'6"N – 4°7'48"W) July 1988, 3 m. We also investigated histologically a piece of the neotype and a piece of Halisarca franz-schulzei Merejkovsky, 1879 from Zoological Institute RAN, St-Petersburg, Russia.
Description. External morphology. Predominantly incrusting sponge, 0.3–12 mm thick, forming smooth crusts on shallow water hard surfaces, or globular, 0.5–2 cm in diameter. Frequently a fouling organism (on red and brown algae, ascidians, shells of bivalves and brachiopods, tubes of sedentary polychaetes, hydroid stalks) ( Figs. 2, 3 View FIGURES 2 – 5 ). Incrusting forms have multiple oscula 0.3–1 mm in diameter, scattered regularly; globular sponges have one osculum or, occasionally, two oscula with raised rims, approximately 1–2 mm in diameter. Pores are dispersed. A significant reduction in the aquiferous system is characteristic for the latest phases of reproduction (embryo and larvae development). During these periods oscula and pores are rarely observed. Colour is yellow-beige to pale brown ( Fig. 2 View FIGURES 2 – 5 ); during last phases of reproduction the specimens have ivory or whitish colour due to the presence of embryos and larvae ( Fig. 3 View FIGURES 2 – 5 ). Surface is smooth and slimy. Consistency is soft, gelatinous and slightly elastic.
Anatomy. Soft tissues. An ectosomal region up to 27 µm thick, consists of three layers: (1) The upper layer composed of a mucous acellular cuticle 0.1–0.2 µm thick ( Figs. 4 View FIGURES 2 – 5 , 6 View FIGURE 6 ), underlying external parts of T-shaped exopinacocytes and a diffuse collagen region 0.9–3 µm thick beneath it; (2) the middle layer ~ 5–10 µm thick containing interlaced collagen fibrils organized into firm tracts. This layer can include rare spherulous cells; and (3) the inner layer, up to 3–7 µm thick, consisting of condensed collagen fibrils and the cell bodies of T-shape exopinacocytes. The choanosome makes up the greatest volume of the sponge body and is composed of choanocyte chambers, canals and mesohyl ( Fig. 4 View FIGURES 2 – 5 ). The organization of the aquiferous system is reminiscent of a syconoid system described earlier ( Lévi 1956; Vacelet et al. 1989). The long, branching, tubular, meandering choanocyte chambers are arranged radially around a large exhalant canal ( Fig. 4 View FIGURES 2 – 5 ). Choanocyte chambers could vary in abundance and size depending on life cycle. During active growth period choanocyte chambers are 120–600 µm long and 24–90 µm in diameter. Abundance and length of choanocyte chambers decrease during reproduction ( Fig. 5 View FIGURES 2 – 5 ).
Cytology. Choanocytes ( Figs. 7, 8 View FIGURES 7 – 15 ) are cylindrical or trapeziform, with long and numerous basal pseudopods ( Vacelet et al. 1989; Boury-Esnault et al. 1990; Gonobobleva & Maldonado 2009). The cells are 6–9.5 μm long and 3.5–5 μm wide, with a smooth flagellum and a collar composed of 30–40 microvilli. Some but not all cells have a periflagellar asymmetrical cytoplasmic sleeve. Choanocytes within a choanocyte chamber are connected with each other at the basal-lateral surfaces with numerous interdigitations. Nuclei are pyriform (2.5 x 4.8 μm) with apical thinning and often with one to three nucleoli. Cytoplasm contains phagosomes, electron transparent vacuoles, perinuclear Golgi complexes, and mitochondria.
Exopinacocytes ( Figs. 9–11 View FIGURES 7 – 15 ) are T-shaped. External flat cytoplasmic parts connect with each other by interdigitations. Cell bodies, including the nucleus and the main cytoplasm volume, are situated beneath the ectosome. Cell bodies are connected with flat cytoplasmic parts by thin and long cytoplasmic bridges. Nuclei are nucleolated.
Endopinacocytes ( Figs. 12, 13 View FIGURES 7 – 15 ) are flat fusiform or polygonal in shape. A nucleolated nucleus is oval, 1.6– 2 x 3–3.8 μm in size, and is located in the center of the cell. The cytoplasm includes well-developed Golgi complexes, small electron dense spherical inclusions and abundant electron-transparent vacuoles.
Basopinacocytes ( Figs. 14, 15 View FIGURES 7 – 15 ) are flattened and polygonal, about 8–14 μm in length and about 2 μm in width. The cytoplasm includes a well-developed Golgi complex, small electron dense spherical inclusions and some phagosomes.
Mesohylar cells. Lévi (1956) was the first to describe the free cells in the mesohyl of H. dujardini at light microscopy level. He distinguished three cells populations (types): (1) amoeboid or star-shaped cells with spherical central nucleus and clear cytoplasm containing small granules, called “amoebocytes I and II”, or amoebocytes and collencytes; (2) the cells with the cytoplasm filled with acidophilic granules, called “amoebocytes III”, or fuchsinophil cells, or granular cells; and (3) cells with large peripheral nucleus and the cytoplasm filled with big spherical inclusions – spherulous cells. In subsequent works ( Bergquist 1996; Bergquist & Cook 2002) three types of mesohylar cells were also mentioned: (1) lophocytes; (2) spherulous cells, common in the zone between ectosome and choanosome and along canals; and (3) fuchsinophilic cells scattered throughout the choanosome. However, the use of electron microscopy led to recognition of six mesohylar cells types in H. dujardini : (1) amoebocytes, (2) collencytes or lophocytes, (3) spherulous cells, (4) granular cells, (5) microgranular cells, and (6) vacuolar cells ( Volkova & Zolotareva 1981; Korotkova & Ermolina 1986; Sukhodolskaya & Krasukevitch, 1984), which are described below.
(1) Archaeocytes (“amoebocytes I”) ( Figs. 16, 17 View FIGURES 16 – 19 ) are abundant cells in H. dujardini mesohyl. These cells have an irregular amoeboid shape and often produce pseudopodia. A large spherical nucleolated nucleus (2.5–3.5 µm in diameter) is located in the center of the cell. Cytoplasm is characterized by the absence of any special or monotypical inclusions, although it can contain heterophagosomes and various osmiophilic or electron-transparent inclusions. There are well developed Golgi complex and endoplasmic reticulum. Archaeocytes are distributed throughout the mesohyl.
(2) Lophocytes ( Figs. 18, 19 View FIGURES 16 – 19 ) have a star-like, clavate, or oval form, often with a tuft of secreted collagen fibrils attached to the lateral and posterior ends. The cytoplasm contains some phagosomes and small osmiophilic inclusions. An oval nucleolated nucleus is located in the anterior part of the cell.
(3) Spherulous cells ( Fig. 20 View FIGURE 20 ) are oval or round in appearance (8–12 µm in diameter) and are characterized by the presence in the cytoplasm of 2–7 large trapeziform, spherical, or oval homogenous osmiophilic inclusions 2–8 µm in diameter. These inclusions have a microgranular structure. The cytosol is reduced to a thin layer between these inclusions. The cytoplasm sometimes contains large heterogenous inclusions and small electron transparent vacuoles. The Golgi complex is well developed. A nucleus without nucleoli could be in the cell’s center or at the periphery. Spherulous cells can be distributed anywhere in the mesohyl, but are more common in external parts of the choanosome. They often occupy the collagen-reinforced layer in the sponge ectosome.
(4) Granular cells (amoebocytes III or fuchsinophilic cells) ( Fig. 20 View FIGURE 20 ) form one of the more numerous cell types in the mesohyl. The shape of these cells is highly variable; usually cells are irregular, ovoid or elongate with a mean diameter about 8 µm. The cytoplasm is filed with round or oval electron-dense homogenous granules (0.5– 2 µm in diameter). The nucleus has a peripheral, or, rarely, central location in the cell. The cells are distributed throughout the choanosome.
(5) Microgranular cells ( Fig. 20 View FIGURE 20 ). We propose to distinguish this cell type in H. dujardini from granular cells. Microgranular cells are characterized by irregular, amoeboid or, rarely, oval shape and have large lobopodia. The cytoplasm is filed with abundant small (0.2–0.5 µm in diameter) electron-dense spherical or oval inclusions. These inclusions can be of oval shape or rod-shaped. The Golgi complex is well developed. The nucleus can be nucleolated or without nucleoli, located in the cell’s center or at the periphery. Microgranular cells do not display any special localization in the choanosome.
(6) Vacuolar cells ( Fig. 20 View FIGURE 20 ) are rare in the mesohyl of H. dujardini but do not represent a stage in the ontogenesis of spherulous cell, as thought by Lévi (1956). Cells are irregular to oval in shape, 5x 7–9x12 µm in size. The cytoplasm contains one to 12 vacuoles of oval or elongated shape 0.5–7.2 µm in diameter. Vacuoles are electrontransparent or could contain very loose mucus-like inclusions, and in sometimes intact symbiotic bacteria. Vacuolar cells are predominantly distributed in the middle part of the choanosome.
Additional transient mesohylar cell types could differentiate at various stages in the life cycle. They can be nutritive cells, macrophages, and special granular cells containing cationic peptides and proteins.
Nutritive cells ( Fig. 21 View FIGURES 21 – 23 ) are a mixed heterogeneous cell population with predominance of dedifferentiated choanocytes. Choanocytes filled with phagosomes leave choanocyte chambers and enter the mesohyl during oocytes vitellogenesis ( Korotkova & Apalkova 1975; Korotkova & Aisenshtadt 1976; Aisenshtadt & Korotkova 1976). Among the nutritive cells there are also amoebocytes I and spherulous cells. Nutritive cells are phagocyted by growing oocytes for yolk granules formation.
Macrophages ( Fig. 22 View FIGURES 21 – 23 ) are large cells that appear at the end of sexual reproduction when begins the postreproductive restoration phase of the life cycle (Ereskovsky 2000), or during the regeneration when it undergoes destructive processes in the mesohyl ( Korotkova & Movchan 1973; Volkova & Zolotareva 1981; Korotkova & Ermolina 1986b; Sukhodolskaya & Krasukevitch 1984). Cell dimensions can reach 30–35 µm, the cytoplasm is filled with large heterogeneous phagosomes. A large spherical nucleus is nucleolated.
Special eosinophilic granular cells ( Fig. 23 View FIGURES 21 – 23 ) contain cationic peptides and proteins ( Krylova et al. 2004). These cells are amoeboid or oval in shape (about 8–12 µm in diameter); the nucleus is oval and voluminous (about 3.2 µm in diameter) with condensed heterochromatin or a small nucleolus. The cytoplasm contains several specific inclusions: (1) vacuoles that include one to ten round or oval electron-dense granules with peripheral thinly-fibrillar material; (2) vacuoles with electron-dense fine-granular inclusions; (3) electron-dense thinly-fibrillar inclusions located in the cytoplasm. Cells differentiate at the beginning of vitellogenesis of oocytes. During the embryonic development they concentrate in the mesohyl surrounding embryos. These eosinophilic granular cells (or eosinophilic amoebocytes) of maternal sponge penetrate developing embryos and remain there until the beginning of metamorphosis ( Korotkova & Ermolina 1982; Ereskovsky & Gonobobleva 2000; Gonobobleva & Ereskovsky 2004a, b).
Symbiotic bacteria. Symbiotic bacteria of H. dujardini are represented by a single morphotype found both in the mesohyl of all investigated adults ( Fig. 20 View FIGURE 20 ) and during all stages of embryonic development of the White Sea specimens ( Ereskovsky et al. 2005). Bacteria are almost evenly distributed in the mesohyl and do not form accumulations. Sometimes bacteria are located in vacuoles of archaeocytes. Bacteria (likely gram-positive) have a curved spiral form, characteristic of spirills ( Fig. 20 View FIGURE 20 ). Their length is about 0.45 µm, thickness 0.18 µm, the cell wall is 0.05 µm thick. No flagella or piles are found at the surface of bacterial wall. The cytoplasm is heterogeneous. Its peripheral part has a medium electronic density, while the central part is electron dense and possibly corresponds to the nucleoid. In-between, a thin electron-transparent layer is visible.
Reproduction. The sponge is dioecious (however some rare individuals could be hermaphrodites) with short seasonal period of reproduction ( Table 2 View TABLE 2 ). Spermatogenesis occurs only during the cold part of the year (Ereskovsky 2000). Larvae are sub-spherical to oval dispherulae, 100–200 µm in diameter, completely ciliated but only sparsely so at the posterior pole ( Ereskovsky & Gonobobleva 2000; Gonobobleva & Ereskovsky 2004a, b).
Habitat. Halisarca dujardini is usually found at depths from 1 to 20 m but also from upper littoral (Barents Sea) to 300 m (Skagerrak), mainly on the algae Fucus vesiculosus , F. serratus, Ascophyllum nodosum and Laminaria saccharina , under and on stones and boulders, in empty shells of lamellibranchs, on carapaces of crabs, at the base of gorgonians, and on the ascidian Styela rustica .
Distribution. Halisarca dujardini species complex is considered to be cosmopolitan ( Bergquist & Cook 2002). In fact, however, its distribution is limited to temperate waters: in the North Atlantic—along the European costs from English Channel to White Sea and in Atlantic coast of USA (Massachusetts) and in the North-West of Mediterranean Sea; in North Pacific—Bering Sea, Avacha Bay and in Russian coast of Japan Sea; in South Pacific—the New Zealand coasts ( Fig. 24 View FIGURE 24 ). See Annex 1 for the detailed distribution.
Material examined. Holotype: collected in “Grotte à Corail” Maire Island, 16 m depth, 43°12'38.04"N – 5°19'56.80"E, Muséum National d'Histoire Naturelle (Paris, France) MNHN DJV 128. Paratype: collected in “Grotte à Corail” Maire Island, 16 m depth, 43°12'38.04"N – 5°19'56.80"E, Zoological Institute RAS (Saint-Petersburg, Russia) № 11141.
Description. External morphology. Thinly incrusting sponge, 0.17–0.19 mm thick, spreading on the surface of the living bryozoan Smittina cervicornis ( Fig. 25, 26 View FIGURES 25, 26 ). The sponge covers the host’s colony from the base to the tips of the branches with a continuous layer. Most often, the sponge coating is very thin and transparent, and thus hardly visible. The sponge may become relatively thick and then appears as an opaque sheath. The surface is skinlike, lustreless and smooth, even ( Fig. 26 View FIGURES 25, 26 ). Oscula have a cone shape (height: 0.1–0.5 µm and diameter about 0.5 mm) and are located exclusively along the bryozoan branch edges that are devoid of zooid orifices. Texture is soft, very delicate and easily torn. The colour is pale-yellow.
Internal organization. Soft tissue organization. There is a thin ectosomal region, 4–14 µm deep, consisting of the flat part of exopinacocytes and of an underlying dense collagen layer, about 1.7–2.6 µm deep ( Fig. 27 View FIGURE 27 – 29 ). This layer contains symbiotic bacteria and some secretory cells ( Fig. 27 View FIGURE 27 – 29 ). Cell bodies of T-shape exopinacocytes and rare spherulous cells occur beneath the collagen layer.
The choanosome makes up most of the volume of the sponge body and is composed of choanocyte chambers and mesohyl ( Figs. 27, 28 View FIGURE 27 – 29 ). The individual choanocyte chambers are tubular in shape, sometimes branched, measuring from 76 up to 175 μm in length and are 9.3–14 μm wide. There are mobile cells present in the mesohyl.
The basal part ( Fig. 29 View FIGURE 27 – 29 ), in contact with the bryozoan surface, includes a dense, thin (0.17 µm) cuticle secreted by the basopinacocytes. Collagen cuticle includes spiral symbiotic bacteria and other bacteria, which are not present in other sponge parts.
Cytology. Exopinacocytes ( Fig. 30 View FIGURES 30 – 35 ) are T-shaped with a thin, flattened external cytoplasmic part, and an internal cell body. The cell body is irregular or triangular in shape, about 4.9x5.3 μm. The two parts of the cell are connected by a thin cytoplasmic bridge. Nucleus is lightly ovoid (2.2–2.4 µm in diameter), often without nucleolus.
Choanocytes ( Fig. 28 View FIGURE 27 – 29 , 31 View FIGURES 30 – 35 ) are elongate or roughly pyramidal, 2.3–5.3 µm wide at the base and 1.9–4.2 µm wide at the nucleus area, the height is 4–5.9 µm. Choanocyte collar arises at approximately 4/5 of the height of the cell and is composed of about 18 microvilli. Nucleus is central or apical, spherical about 1.9 µm in diameter, occasionally with a nucleolus, 0.5 µm in diameter. The apical part of choanocytes encompasses many small vacuoles. The central and basal cytoplasm usually contains 2–4 phagosomes, 0.8–1.5 µm in diameter with heterogeneous inclusions. The choanocytes also contain mitochondria, osmiophilic granules (0.3–0.6 µm), electron transparent vacuoles (0.1–0.8 µm) and other inclusions. The choanocytes contact each other only at the base. Long fine pseudopodia frequently arise from the central and basal parts of the choanocytes; the latter anchor choanocytes in the mesohyl. The free apical surface of the choanocytes and the flagella are covered by a thin (1 µm) glycocalyx layer.
Endopinacocytes ( Fig. 32 View FIGURES 30 – 35 ) are flat cells with very thin, long, and flat cytoplasmic outgrowths (about 0.2–0.3 µm in thickness), non-flagellated, 9–13.7 µm wide by 2.1–2.5 µm high in the nucleus area. The cytoplasmic outgrowths can make branching diverticules. Endopinacocytes are anchored in the mesohyl by short, thin basal pseudopodia, which develop from the central part of the cell. Nucleus is ovoid (2 x 2.3 µm), often without nucleolus. The cytoplasm usually contains 1–3 phagosomes 0.4–0.6 µm in diameter, transparent vacuoles, and osmiophilic inclusions.
Basopinacocytes ( Fig. 33 View FIGURES 30 – 35 ). These cells constitute the loose layer underlying the base of the sponge. They are oval, flattened in basal part, about 5.7 μm in length and about 1.8 μm in width in the nucleus region. The nucleus is oval 0.9 x 2.8 μm in size. The cytoplasm includes a well-developed Golgi complex, small electron dense spherical inclusions and abundant mitochondria.
Mesohylar cells. Archaeocytes ( Figs. 29 View FIGURE 27 – 29 , 34 View FIGURES 30 – 35 ) are amoeboid cells, 5– 7 x 4–2.9 µm in size with a round, nucleolate nucleus (1.9–2.2 µm in diameter, nucleolus: 0.5 µm). The cytoplasm contains phagosomes (0.5–2 µm in diameter) with heterogeneous inclusions, round electron dense inclusions (0.2–0.7 µm in diameter) and, sometime, lipid droplets 0.5–0.7 µm in diameter. Archaeocytes are not abundant and are dispersed in all parts of the sponge mesohyl.
Lophocytes ( Fig. 35 View FIGURES 30 – 35 ). Clavate, drop-like or oval cells (6.2–8.3 x 1.3–2.4 µm), often with a tuft of secreted collagen fibrils attached to the posterior end. The nucleus, located in the anterior part of cell, is oval (about 1.4 x 2.7 µm) with a nucleolus 0.5 µm in diameter. Cytoplasm contains mitochondria, some digestive vacuoles, and small osmiophilic inclusions.
Spherulous cells ( Figs. 28 View FIGURE 27 – 29 , 36 View FIGURES 36 – 43 ). Oval, round, or irregular in shape cells, 7.9–11.2 µm in diameter. Nucleus is round (about 3.4 µm in diameter) or oval, commonly without the nucleolus. The cytoplasm contains large, trapeziform or, more rarely, spherical (0.6–2.9 µm in diameter) osmiophilic inclusions. The Golgi complex is well developed. Spherulous cells are more abundant in the exterior part of the choanosome and in the ectosome, but they are also present in the choanosome with a preferential distribution under the choanocyte layer.
Vacuolar cells ( Fig. 29 View FIGURE 27 – 29 , 37 View FIGURES 36 – 43 ) with big electron transparent vacuoles are often situated in the basal part of sponge choanosome. Cell form is irregular, sometimes dumbbell-like, 5.4–11.7 µm in length and 3.5–4.6 µm in width, with an oval, anucleolate nucleus, in the central part 2x 2.4 µm in dimension. The cytoplasm is reduced to a fine layer surrounding the vacuoles and the nucleus. The vacuoles are oval or irregular, from 2.5x4.2 to 4 to 6.8 µm in size.
Microgranular cells ( Fig. 38 View FIGURES 36 – 43 ) are large amoeboid cells, 5.7 x 7.4 µm. The nucleus is round (2.7 µm in diameter) with a nucleolus 0.4 µm. The cytoplasm is filled by oval (length 0.5–1.15 µm; diameter 0.15–0.45 µm) or round (0.2–0.4 µm in diameter) electron-dense granules, surrounded by a membrane and by abundant small irregular, electron transparent vacuoles.
Granular cells ( Fig. 39 View FIGURES 36 – 43 ) are oval or irregular cells with thin cytoplasmic outgrowths. Cell length is 4.1–5.6 µm and width is 2.7–3.6 µm. The anucleolate nucleus is round or ovoid, about 1.7 µm in diameter. The cytoplasm is filled by oval or round electron-dense homogenous granules 0.2–1.5 µm in diameter. The granules are surrounded by a thin electron transparent space.
Rhabdiferous cells ( Fig. 40 View FIGURES 36 – 43 ) are amoeboid in shape (6.2– 11 x 2.7–6.1 µm) with long pseudopodia. The nucleus is oval, about 2 µm in diameter with a nucleolus 0.4 µm. The cytoplasm is filled with rhabdite-like or oval osmiophilic inclusions 0.5–2.3 µm in length and 0.2–0.9 µm in diameter.
Symbiotic bacteria. Two morphotypes of symbiotic bacteria, B1 and B2, occur in the mesohyl. Both are extracellular. The type B1 ( Figs. 41, 42 View FIGURES 36 – 43 ) is found in the mesohyl as well as in the surface layer of the sponge. These bacterial cells are spiral-shaped, slightly curved, with flagella 0.24–0.31 µm in diameter and 2–4.5 µm in length. The cytoplasm is electron-dense. Thick nucleoid filaments form a regular, thin central structure. The cell wall is surrounded by a thin wavy extracellular layer. An electron-dense thickening of the cell wall forms a spiral around the cell surface. The type B2 ( Fig. 43 View FIGURES 36 – 43 ) is located only in the basal collagen layer lining the border with the bryozoan host. These bacteria are oval, 0.8–1.2 µm in length and 0.4–0.5 µm in diameter. The cytoplasm is homogeneous and of medium electron density. The nucleoid zone contains a filamentous, irregular network which appears characteristic of these bacteria. Most remarkable are numerous thin projections, up to 0.6 µm long, that extend from the surface of the bacteria.
Mitochondrial genome. Mitochondrial genome of H. harmelini is identical in both the gene content and the gene arrangement to those of H. dujardini and Chondrilla nucula (Demospongiae, Chondrosida ) (Fig. 44). Most of the protein genes and the structural RNA genes are identical or similar in size between the two Halisarca genomes ( Table 3 View TABLE 3 ), but the total length of intergenic regions is larger by ~ 1,300 bp (6.8% of the genome size) in H. harmelini . Comparison of coding sequences among H. harmelini , H. dujardini , and C. nucula revealed that genetic distances between the two Halisarca species (on average ~ 0.12 for all sites, 0.53 for 4-fold degenerate sites, see Table 3 View TABLE 3 for individual proteins) are similar to genetic distances between either of them and C. nucula ( Fig. 45 View FIGURE 45 ). The extent of sequence divergence between the two species greatly exceeds the amount of differences found between two specimens of H. dujardini and within closely related C. nucula ( Fig. 45 View FIGURE 45 ), sampled across a large geographic range and various habitats ( Duran & Rützler 2006). Given that the rate of sequence evolution is very low in sponges ( Lavrov 2010), these large genetic distances indicate an ancient divergence between the two Halisarca species. Nevertheless, they still form a strongly supported clade in demosponge phylogeny based on mitochondrial DNA data ( Fig. 46 View FIGURE 46 ).
FIGURE 44. Genomic maps of Halisarca harmelini and Halisarca dujardini mtDNA. Protein (blue) and ribosomal genes (green) are atp6, atp8–9: subunits 6, 8 and 9 of F0 adenosine triphosphatase (ATP) synthase; cox1–3: cytochrome c oxidase subunits 1–3; cob: apocytochrome b; nad1–6 and nad4L: NADH dehydrogenase subunits 1–6 and 4L; rns and rnl: small and large subunit rRNAs. tRNA genes (black) are identified by the one-letter code for their corresponding amino acids. Two arginine, isoleucine, leucine, and serine tRNA genes are differentiated by subscripts with trnR(ucg) marked as R1, trnR(ucu) —as R2, trnI(gau) —as I1, trnI(cau) —as I2, trnL(uag) —as L1, trnL(uaa) as L2, trnS(ucu) —as S1, and trnS(uga) —as S2. Genes for inferred initiator and elongator tRNA(Met) are marked as Mf and Me, respectively. Overlaps between adjacent genes are shown in yellow. All genes are transcribed clockwise.
# nucleotides ab Rates of sequence evolution between HH and HD Predicted initiation and termi-
nation codons in HH and HD c a Data for Halisarca dujardini are from ( Wang & Lavrov 2008). b Bold numbers indicate sequences of different lengths comparing to their homologues in Halisarca harmelini . Asterisks denote internal indels. c Identical initiation/termination codons in H. harmelini and H. dujardini are shown only once.
Ecology and distribution. Specimens of Halisarca harmelini sp. nov. occur only as thin sheets on the rigid erect adeoniform ascophoran bryozoan Smittina cervicornis ( Figs. 25, 26 View FIGURES 25, 26 ). The species has been found in all investigated sites along the Northwestern Mediterranean coast (from 43°46'26"N – 7°30'56"E to 42°28'46"N – 3° 9'10"E) ( Fig. 1 View FIGURE 1 ) 14–65 m depth, on coralligenous rocks and detritical sand. It has not been observed on all other large rigidly erect bryozoans living in the same microhabitats.
Etymology. The species name refers to its first collector, Dr. Jean-George Harmelin, Centre d’Océanologie de Marseille, Station marine d’Endoume, Marseille, France.
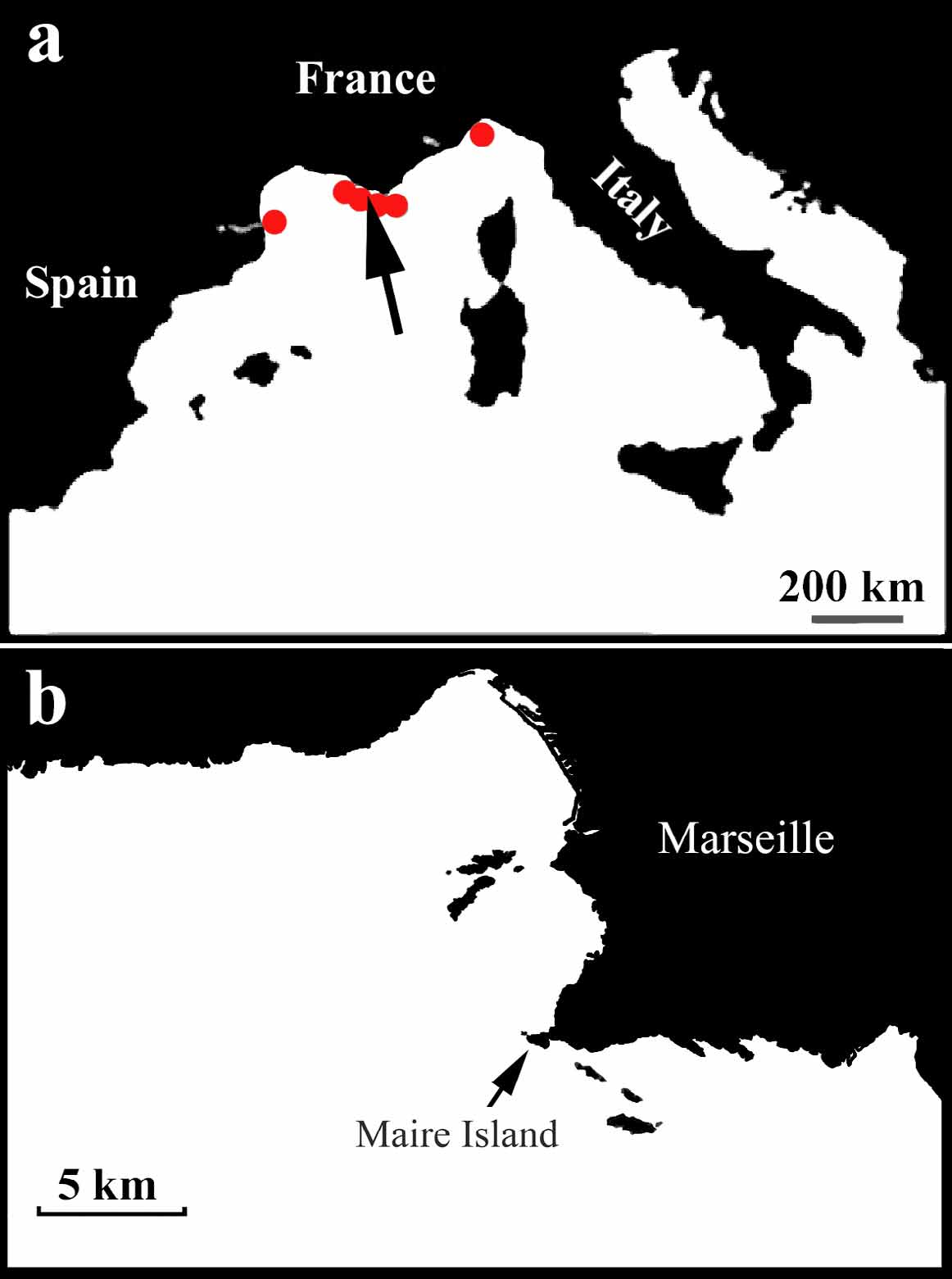
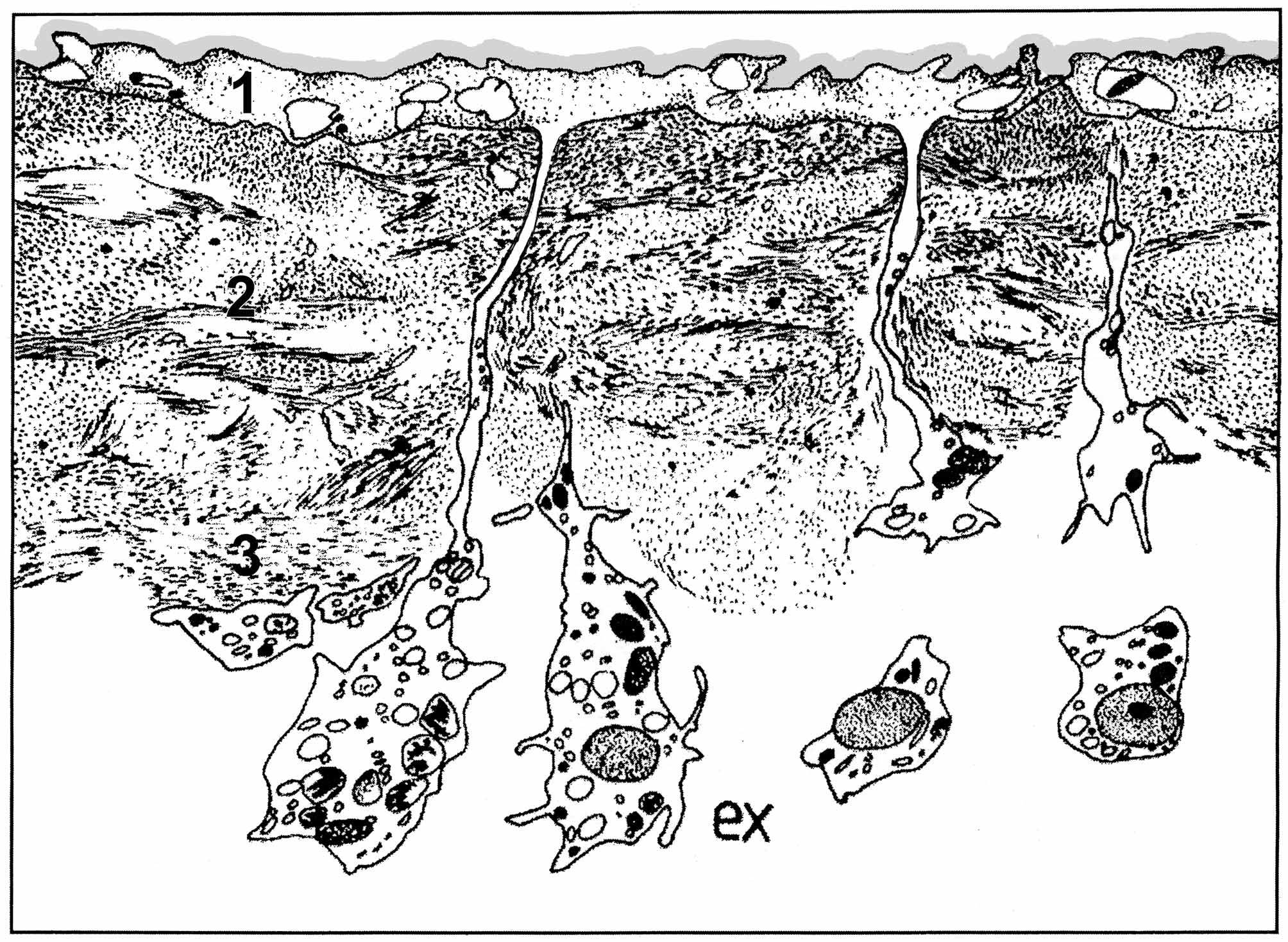
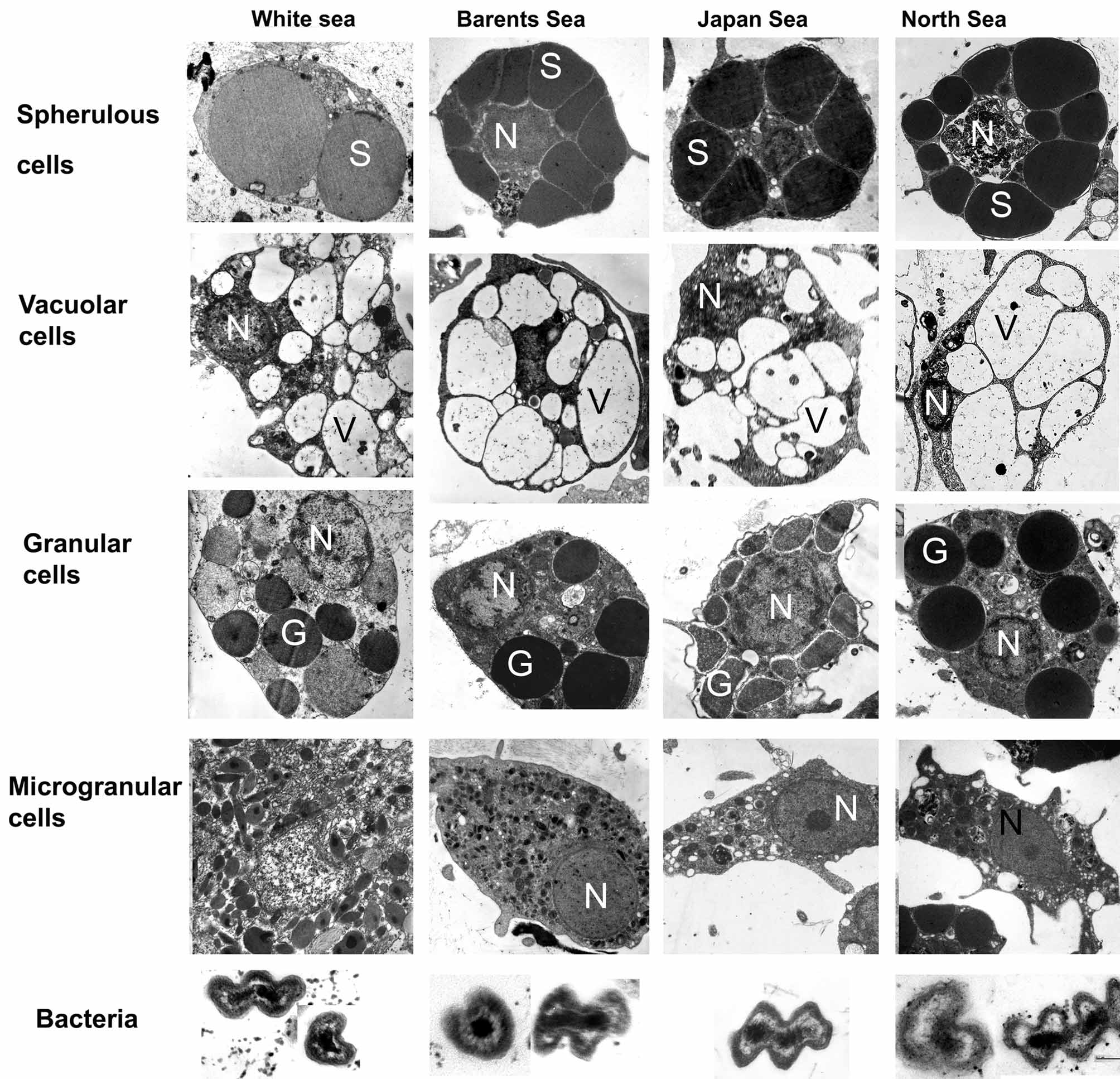
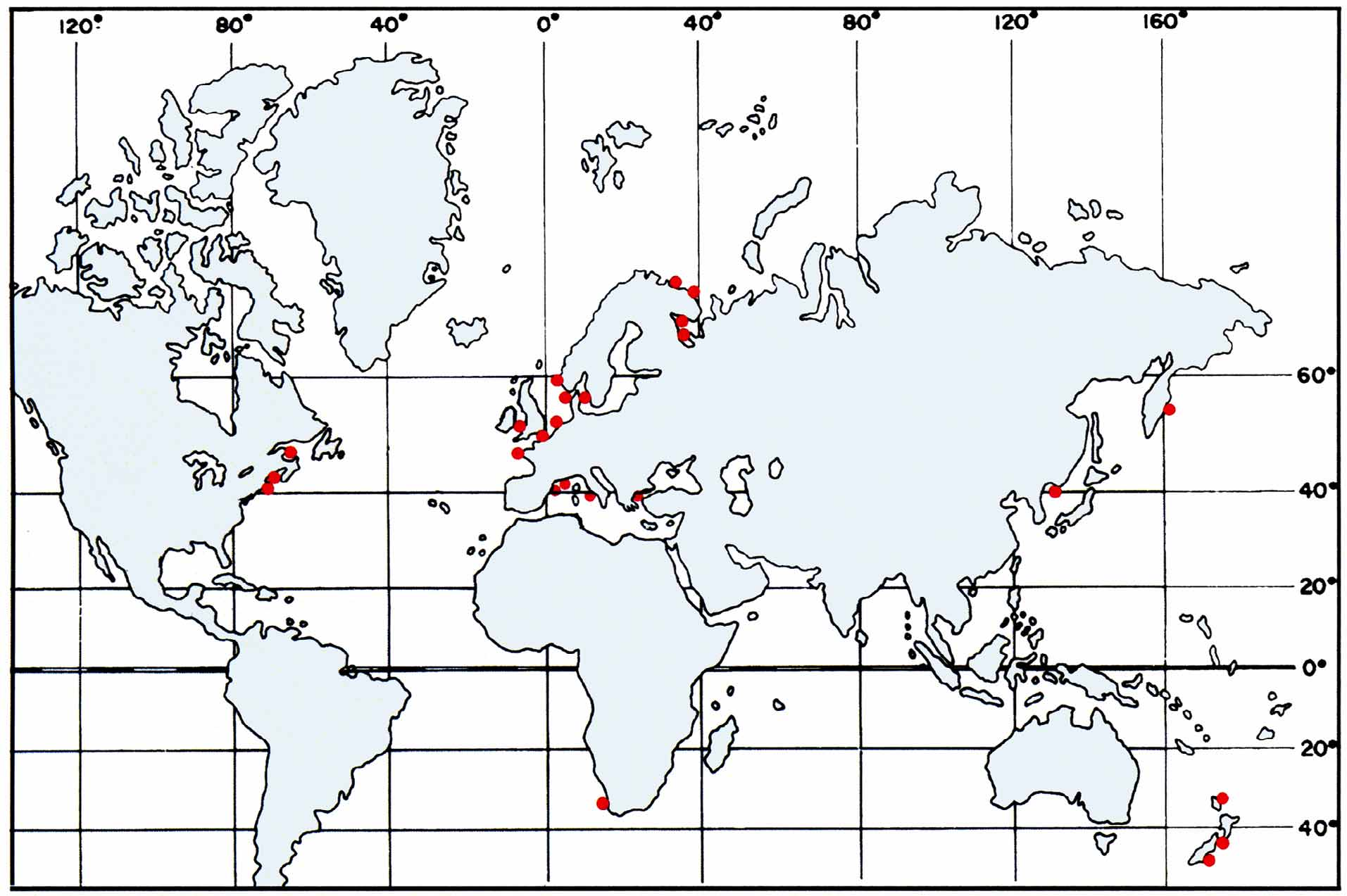
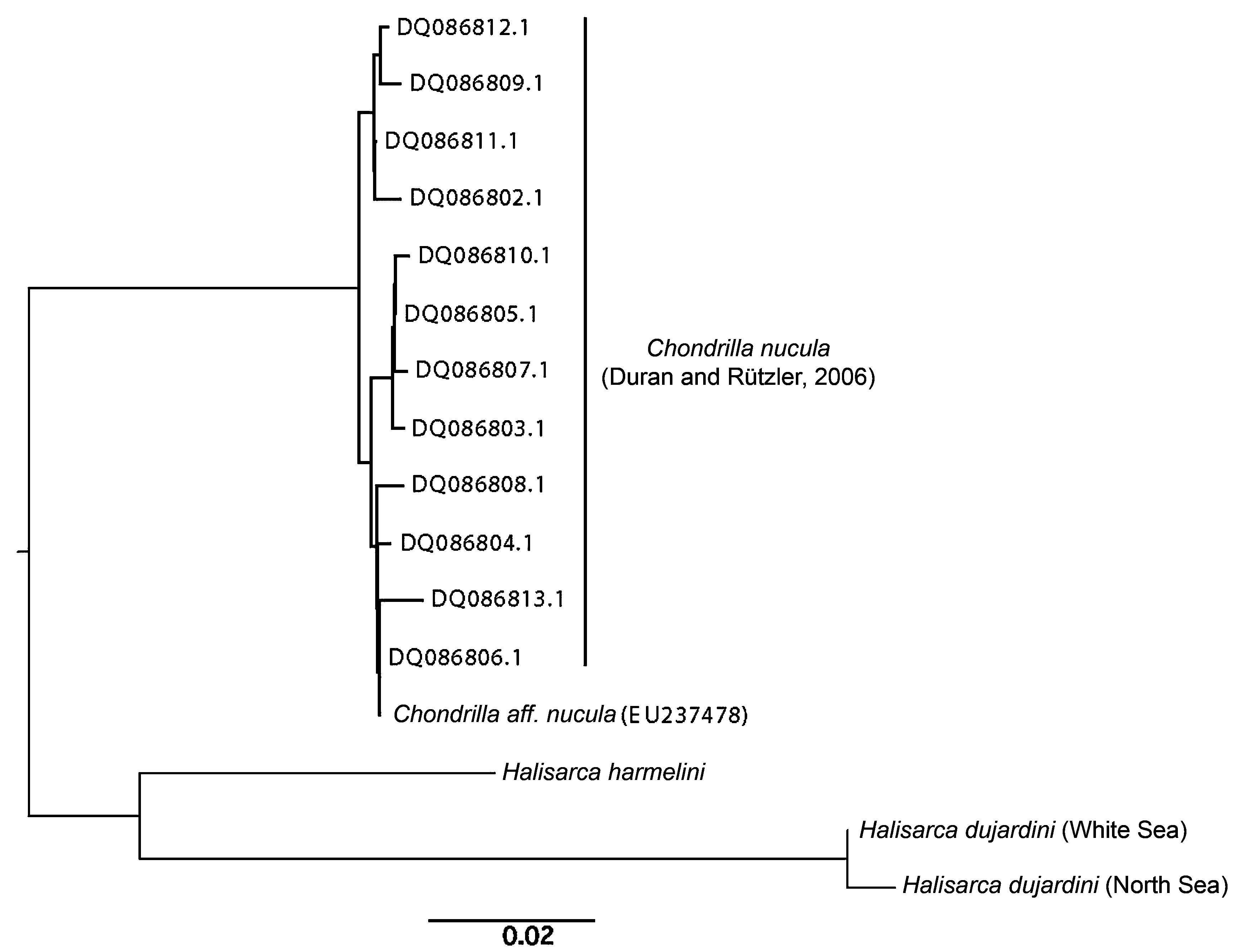
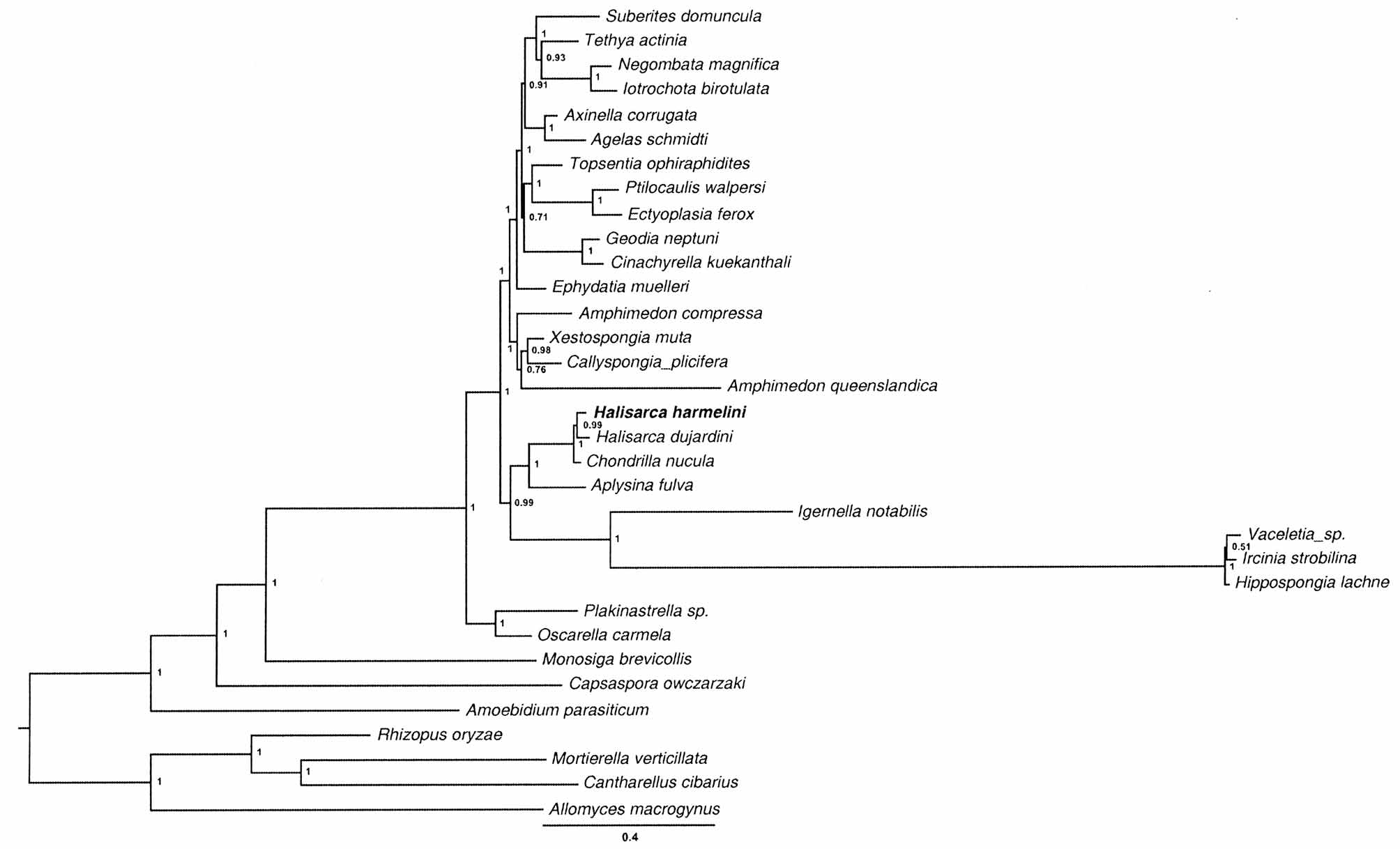
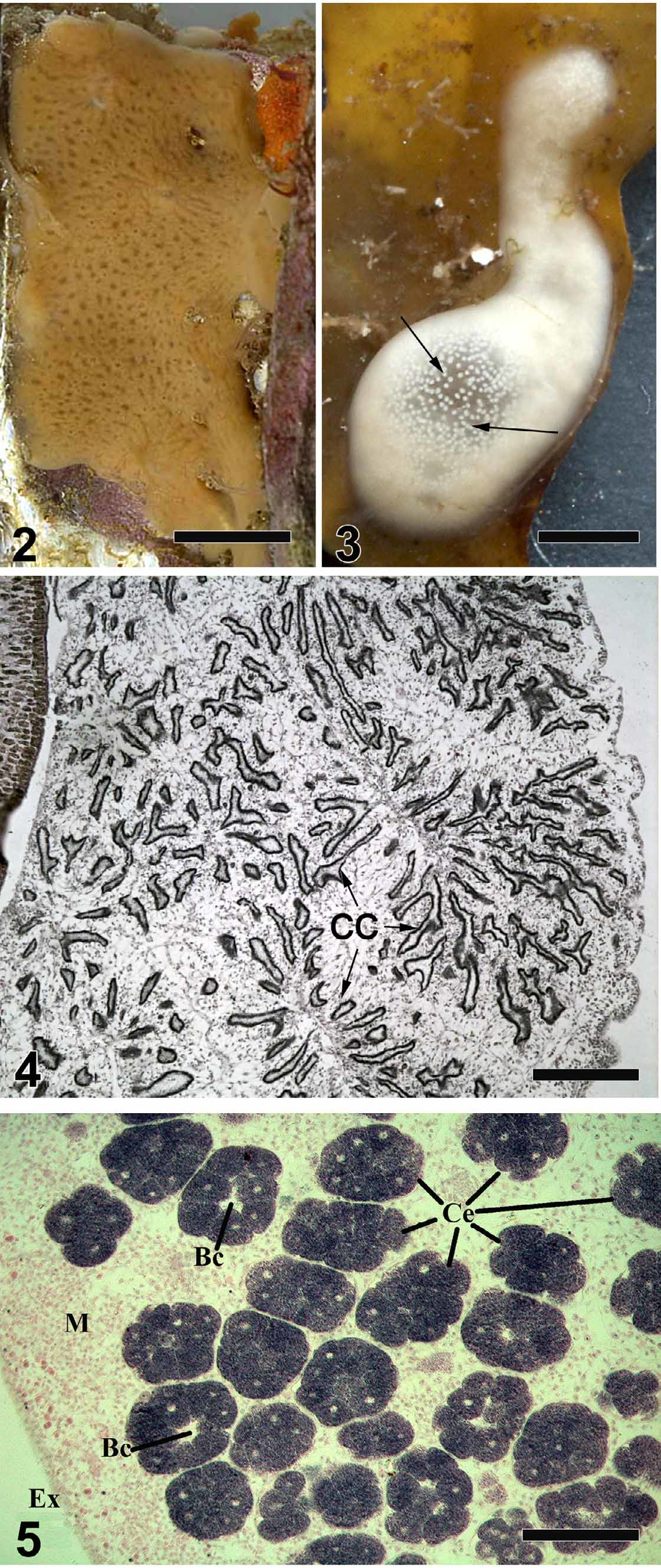
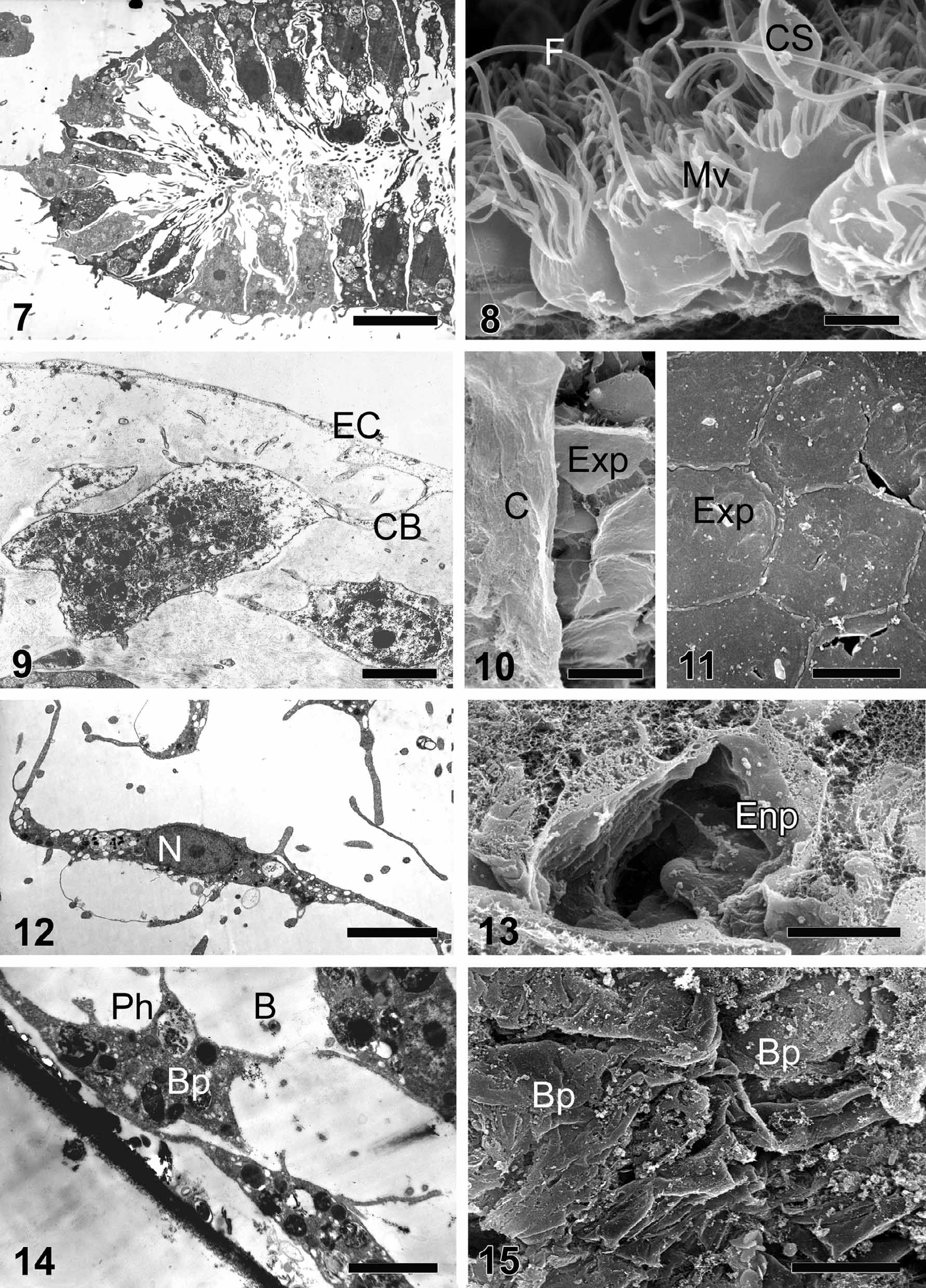
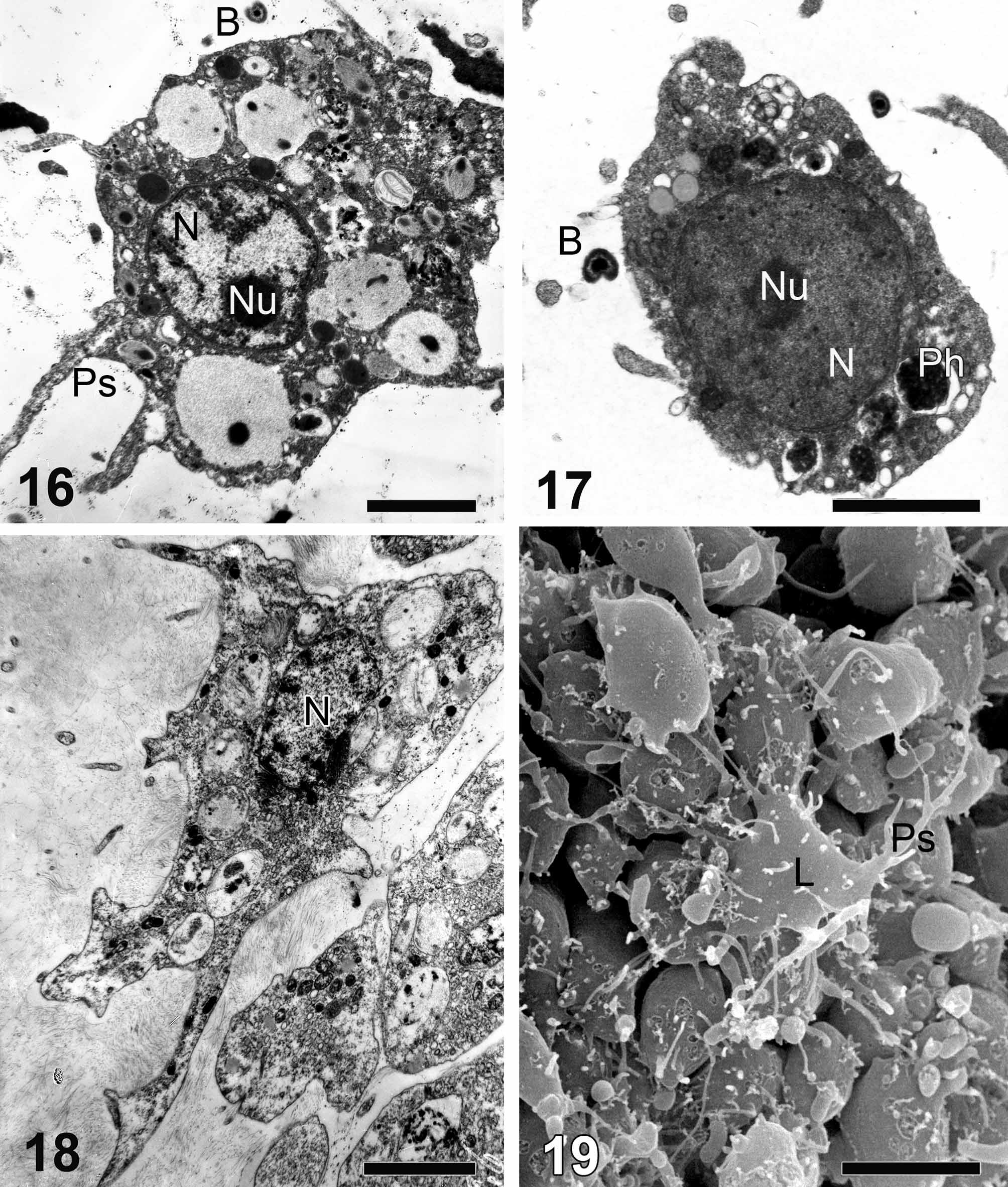

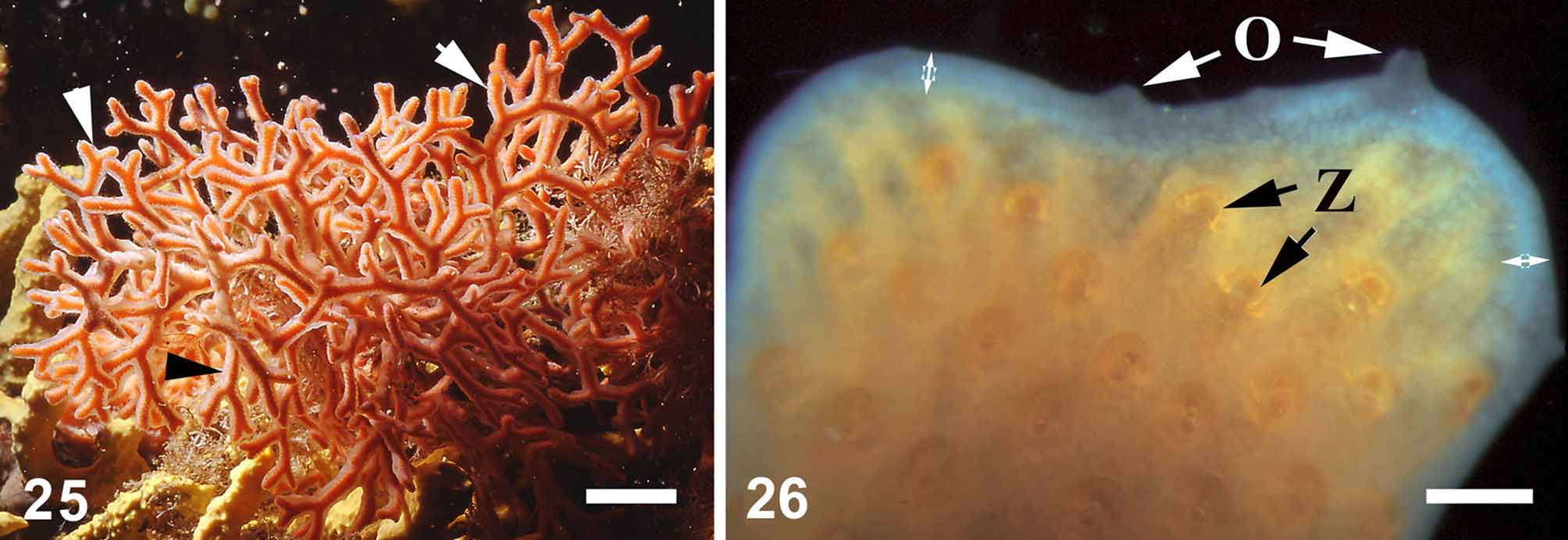

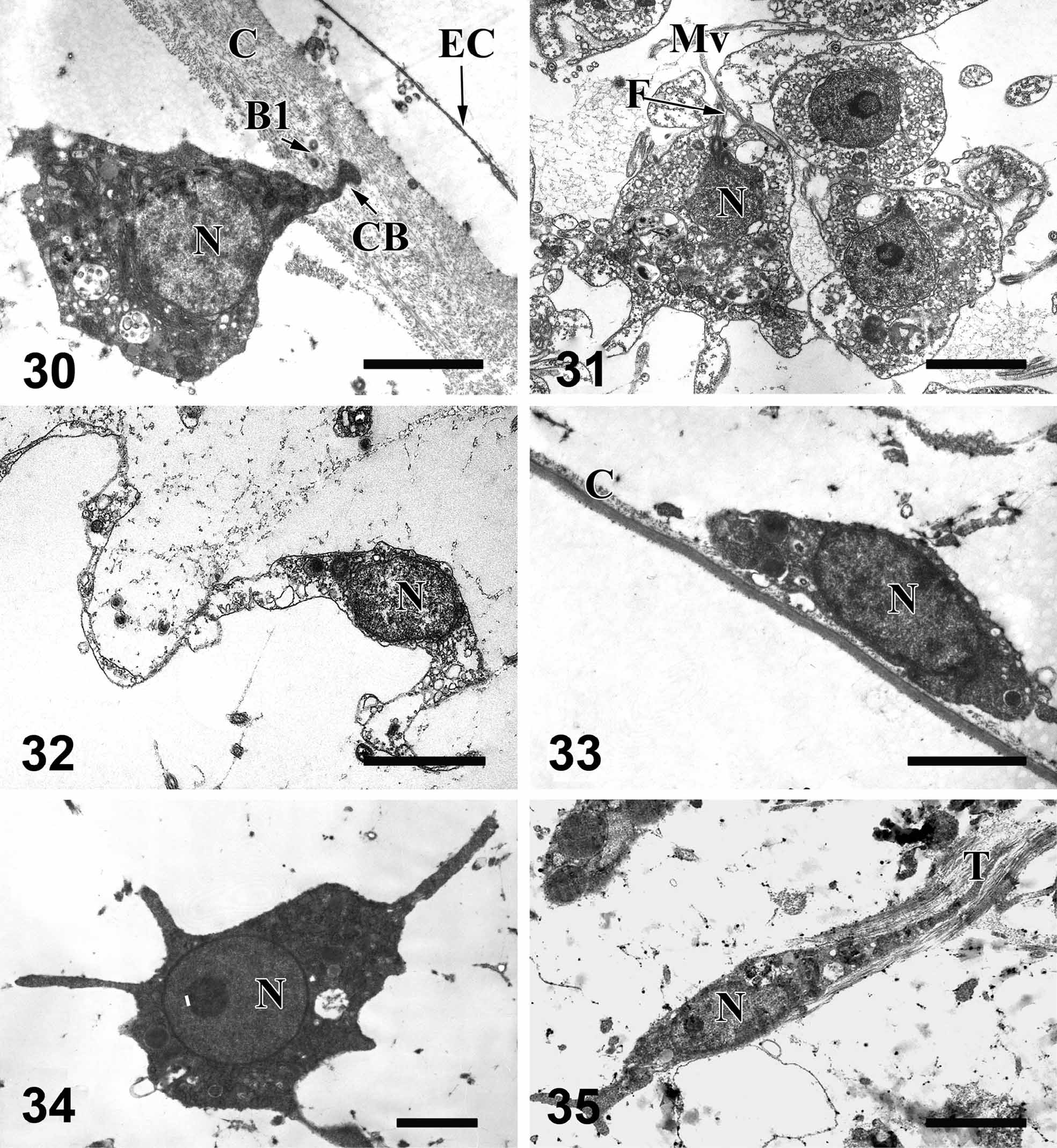

TABLE 2. Period of reproduction (embryonic development and larvae release) in Halisarca dujardini from different geographical regions.
| Region White Sea | Months End of June-July | Ref Ereskovsky 2000 |
|---|---|---|
| Barents Sea North Sea (Bergen) English Channel (Roscoff) SW Pacific (New Zealand) | July-August June-July June-September October-November | Korotkova & Apalkova 1975 Personal observations Lévi 1956 Bergquist 1996 |
| Halisarca harmelini sp. nov. (Figs. 25–46). |
TABLE 3. Comparison of mitochondrial coding and rRNA sequences in Halisarca harmelini sp. nov. (HH) and Halisarca dujardini (HD).
| Gene | HH | HD | All positions dN | dS | dN/dS | Initiation codons | Stop Codons |
|---|---|---|---|---|---|---|---|
| atp6 | 735 | 735 | 0.0905 0.0124 | 0.5593 | 0.0327 | ATG | TAG |
| atp8 | 222 | 225* | 0.1391 0.0587 | 0.7465 | 0.0221 | ATG | TAG/TAA |
| atp9 | 237 | 237 | 0.0834 0.0005 | 0.4871 | 0.0787 | ATG | TAG |
| cob | 1146 | 1146 | 0.1349 0.0388 | 0.9105 | 0.001 | ATG | TAG |
| cox1 | 1563 | 1563 | 0.1236 0.0144 | 1.1801 | 0.0426 | ATG | TAA/TAG |
| cox2 | 771 | 771 | 0.1066 0.0147 | 0.7751 | 0.0122 | ATG | TAG |
| cox3 | 786 | 786 | 0.1035 0.0168 | 0.6306 | 0.019 | ATG | TAA |
| nad1 | 975 | 975 | 0.1418 0.0171 | 1.0451 | 0.0267 | ATG | TAA |
| nad2 | 1425 | 1428* | 0.1581 0.0523 | 1.0367 | 0.0164 | ATG | TAG |
| nad3 | 360 | 360 | 0.1386 0.051 | 1.0314 | 0.0504 | ATG | TAA/TAG |
| nad4 | 1458 | 1470* | 0.1337 0.0008 | 0.7713 | 0.0495 | ATG | TAA |
| nad4L | 300 | 300 | 0.0993 0.0219 | 1.0811 | 0.001 | ATG | TAA/TAG |
| nad5 | 1827 | 1827 | 0.1221 0.0375 | 0.8358 | 0.0203 | ATG | TAA/TAG |
| nad6 | 576 | 591* | 0.1096 0.0315 | 1.0601 | 0.0449 | GTG | TAG/TAA |
| MNHN |
Museum National d'Histoire Naturelle |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Halisarca dujardini
| Ereskovsky, Alexander V., Lavrov, Dennis V., Boury-Esnault, Nicole & Vacelet, Jean 2011 |
Halisarca sputum
| Topsent 1893: 38 |
Hymeniacidon dujardini
| Bowerbank 1866: 224 |
Halisarca dujardini
| Johnston 1842 |
Halisarca dujardini
| Johnston 1842: 192 |