
Simulium (Trichodagmia) chimguazaense, Diaz, Sofia A., Moncada, Ligia I., Murcia, Carlos H., Lotta, Ingrid A., Matta, Nubia E. & Adler, Peter H., 2015
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3914.5.3 |
|
publication LSID |
lsid:zoobank.org:pub:9CA00909-16CC-43F8-BFC0-B15540052EF9 |
|
DOI |
https://doi.org/10.5281/zenodo.5687539 |
|
persistent identifier |
https://treatment.plazi.org/id/A6FBBABF-35AF-498B-B57A-4589405E08A3 |
|
taxon LSID |
lsid:zoobank.org:act:A6FBBABF-35AF-498B-B57A-4589405E08A3 |
|
treatment provided by |
Plazi |
|
scientific name |
Simulium (Trichodagmia) chimguazaense |
| status |
sp. nov. |
Simulium (Trichodagmia) chimguazaense new species
( Figs. 1–13 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 8 View FIGURE 9 View FIGURE 10 View FIGURE 11 View FIGURE 12 )
FEMALE. Total length 3.1–4.3 mm, mean 4.0 mm (n = 5). General body color brownish, with brownish and golden hair.
Head— Frons and clypeus dark brown, with brown hair and with extent of silver pollinosity varying with angle of light; frons nearly square; head 3.3–3.5 times wider than frons. Eyes grayish brown. Maxillary palp slender, brown; palpomere III about 3.2–3.7 times longer than wide, and about 2.8–3.1 times longer than sensory vesicle; neck of sensory vesicle about 1/3 length of its body; palpomere V about 3.9 times longer than III. Antenna dark brown, 0.63–0.73 mm long, lacking long, dense hair (contrary to condition in male), with 9 flagellomeres; pedicel and first flagellomere subequal in size. Lacinia with 26–29 teeth. Mandible with 30–33 short, stout teeth on inner margin and 7–9 thinner, longer teeth on outer margin. Cibarium broadly concave, shallow, without teeth.
Thorax— Length 1.1–1.8 mm. Scutum in dorsal view blackish, with grayish cast and 2 silvery spots anteriorly; in anterior view blackish medially, bordered laterally with broad longitudinal grayish pollinose stripes contiguous anteriorly and extended posteromedially for short distance; silvery spots in anterodorsal view blackish; humeral calli brownish, each with 2 small, silvery patches; scutal hair golden. Scutellum pale brownish, with mixed dark brown and golden hairs. Postnotum dark brown, pollinose. Katepisternum brown, with silvery pollinosity. Anepisternal membrane bare. Wing length 4.0– 4.7 mm; basal section of radius with double row of dark brown setae; stem vein and base of costa with dark brown hair. Halter whitish, with base of stem brownish. Legs brown, slightly paler on basal ¾ of femora and tibiae; basal 1/3 of middle basitarsus and of hind tarsomere I grayish; basal ½ or more of hind basitarsus yellowish white; all hair golden, except brown on hind coxa. Claws each with welldeveloped sub-basal tooth.
Abdomen— Brown dorsolaterally, with pale, pollinose, median, longitudinal stripe on segments I–VII (widest on I and VII); tergites VIII–X grayish pollinose. Pale grayish ventrally, with brownish patches laterally on segments III–IX, shifting to silvery pollinose with anterior lighting. Basal fringe of first abdominal segment with long brown hair, except golden ventrolaterally; remaining segments with brown hair. Sternite VIII bare, with heavily sclerotized, brown, U-shaped concavity anteromedial to hypogynial valves. Cercus in lateral view subquadrangular, rounded posteriorly. Anal lobe ( Fig. 1 View FIGURE 1 B) in lateral view consisting of sclerotized, brown, subrectangular plate with membranous, unpigmented rounded, ventral, setose extension. Hypogynial valve ( Fig. 1 View FIGURE 1 C) small, slender, fingerlike, weakly sclerotized, nearly unpigmented, covered with microtrichia. Genital fork ( Fig. 1 View FIGURE 1 A) slender; stem 1.6–2.3 times longer than arm; lateral plate subrectangular, with long, slender, anteriorly directed apodeme. Spermatheca 1.0–1.2 times longer than wide, darkly pigmented, 43–48% length of stem of genital fork; surface externally with faint polygonal pattern, internally with rows of short spicules; junction with spermathecal duct forming unpigmented area about 25–30% of greatest diameter of spermatheca.
MALE. Total length 4.1–4.7 mm, mean 4.3 mm (n = 4). General body color blackish, with golden and brownish to brassy hair.
Head —Frons and postocciput with long, blackish hair. Clypeus with long, dense, blackish hair swept upward to scape. Antenna with scape bearing long, dense, blackish hair extended at least half length of entire antenna; first flagellomere longest, twice as long as wide; flagellomeres 2–9 slender. Maxillary palp with palpomeres II and III bearing long, blackish hair extended to palpomere IV; sensory vesicle about ¼ length of its palpomere.
Thorax —Scutum blackish, velvety, unpatterned at all angles of light, except slight silvery pollinosity at humeral angles; scutal hair golden, recumbent, uniformly distributed, except denser at humeral angles. Scutellum brown, paler posteriorly, with brassy hair intermixed with few golden hairs. Postnotum dark brown, without hair. Wing length 4.1–4.4 mm; basal section of radius with 1 or 2 rows of brownish setae; stem vein and basicosta with brassy hair. Halter whitish, with stem brownish. Legs brownish, with coxae, apices of femora and tibiae, and entire fore- and midbasitarsi and eutarsi slightly darker, except basal 1/3 of hind basitarsus tan; hair of legs brassy, though mostly golden on femora. Hind tibia spindle-shaped, broadest at 2/3 its length. Hind basistarsus widened, flattened, about 3.5 times longer than wide. Calcipala less than 1/3 width of apex of its segment; pedisulcus about ½ depth of tarsomere I.
Abdomen— Tergites and sternites dark brownish; pleural region brownish, grading to grayish ventrally; tergites II, VI, and VII with dorsolateral pair of silvery spots. Basal fringe and all other abdominal hair brownish to brassy. Genitalia: Gonocoxite 1.2 times longer than wide; outer apodeme markedly extended anteriorly. Gonostylus in ventral view ( Fig. 1 View FIGURE 1 D) 3.1–3.7 times longer than greatest width, smoothly and gradually curved toward apex, about 1.2 times longer than gonocoxite, with 1 small subapical spinule; in lateral view ( Fig. 1 View FIGURE 1 E) slightly sinuous, about 3 times wider basally than subapically. Ventral plate in ventral view ( Fig. 1 View FIGURE 1 D) subrectangular, 1.9–2.0 times wider than long, with hirsute, rounded protuberance centrally in slight concavity of posterior margin; anterior margin convex. Median sclerite, parallel sided, 1.2 times as long as wide, truncate at attachment with ventral plate, bifurcated in apical 1/3. Paramere slender, narrowly connected to series of small, weak spines. Aedeagal membrane with comblike clusters of microsetae.
PUPA. Total length (without gill) 4.3–5.0, mean 4.7 mm (n = 40).
Head —Cuticle smooth. Frontoclypeus bearing 2 trichomes laterally, each with 6 or 7 branches.
Thorax —Cuticle smooth, but with minute rounded and pointed microgranules posteriorly ( Fig. 2 View FIGURE 2 E); 9 trichomes dorsally, each with 10–12 branches, and 2 additional trichomes, each with 3–9 branches at each gill base; pair of trichomes with 4 or 5 branches on each wing sheath. Gill ( Fig. 2 View FIGURE 2 D; Table 2): Length 0.9–1.1 mm, mean 1.0 mm, with 10–12 short, stout, brown to dark gray, transversely furrowed filaments in compact bundle grouped dorsal to ventral as (3 or 4) + 2 + (4 to 6); all branched within basal half of gill, except 1 or 2 pairs of filaments occasionally branched apically; middle pair of filaments thickest; in dorsal view, all filaments in 1 vertical plane, except middle pair directed medially ( Fig. 2 View FIGURE 2 B); transverse furrows shallow, obscure basally, deep, conspicuous in distal 2/3 of gill ( Figs 2 View FIGURE 2 C).
Filaments
12:12 12:11 11:11 11:10 10:10 Individuals 7 2 5 13 17 Abdomen —Tergite I pigmented; all other tergites unpigmented; II with 1 minute, fine seta, 1 minute, short, stout seta, and 4 anteriorly directed, stout setae per side; III and IV each with 1 minute, short, stout seta, and 4 anteriorly directed, recurved hooks per side (largest on IV); V with 2–4 minute, short, stout setae per side; VI with 2 minute, short, thin setae per side; VII with 1 minute, short, thin seta, and small cluster of spine combs ( Fig. 3 View FIGURE 3 D) along anterior margin on each side; VIII with small cluster of spine combs and teeth ( Fig. 3 View FIGURE 3 C) along anterior margin on each side; IX lacking terminal spines. All sternites with clusters of microspines; sternites III–V with 1 or 2 minute, short setae per side; V–VII with pair of stout, anteriorly directed hooks per side ( Fig. 3 View FIGURE 3 B); IX with 2 pigmented plates posteroventrally ( Fig. 3 View FIGURE 3 A). Pleural region of segments VII and VIII each with 1 minute, anteriorly directed seta per side.
Cocoon —Total length along dorsal midline 5.1–6.6 mm, mean 5.8 mm. Boot shaped, tightly and uniformly woven ( Fig. 2 View FIGURE 2 A); individual threads not apparent ( Fig. 2 View FIGURE 2 A, inset); in lateral view, anterior margin only slightly higher than posterior margin of opening.
LARVA (mature) Length 8.1–9.3 mm (n = 6). General body color dark brownish gray, darkest on thorax, palest on abdominal segments V–IX; intersegmental bands thin, less densely pigmented.
Head —Frontoclypeal apotome ( Fig. 4 View FIGURE 4 A) brownish orange, paler than chestnut brown of remainder of head capsule; head spots weakly defined; anteromedial head spots positive, others negative; microsetae sparse; area around ocelli unpigmented. Cervical sclerites ovoid, free from postocciput. Antenna subequal in length to stem of labral fan, uniformly concolorous with frontoclypeal apotome; ratio of articles (proximal to distal including apical sensillum) approximately 1.0:2.0:1.0. Labral fan stout, with 38–49 primary rays. Mandible with sensillum bifid, well developed. Hypostoma ( Fig. 4 View FIGURE 4 C) with slightly concave to straight anterior margin; median and lateral teeth subequal in prominence, extended anteriorly to same level; 3–6 lateral serrations per side; 17–19 unbranched or apically bifurcated hypostomal setae laterally, plus 1 or 2 additional, smaller, unbranched setae per side and 3 or 4 small, unbranched setae on central disk. Postgenal cleft ( Fig. 4 View FIGURE 4 B) miter shaped, about 1.3 times longer than greatest width; extended about 80% of distance from posterior margin of tentorial pits to hypostomal groove; subesophageal ganglion and epidermis within postgenal cleft variably pigmented.
Thorax —Prothoracic proleg with lateral plate weakly sclerotized, pale yellowish brown, small, subrectangular. Gill histoblast of 10–12 short, stout filaments.
Abdomen —Expanded gradually posteriorly, abruptly narrowed at segment VIII; ventral tubercles absent. Cuticle nearly bare. Anal sclerite with anterior arms broad; posterior arms slender, and about 1.8 times longer than anterior arms. Rectal papillae of 3 lobes with more than 25 fine lobules each. Posterior circlet with 200–221 rows of 26–32 hooklets each.
Polytene chromosomes. The polytene complements (2 n = 6) of 36 larvae (15 females, 21 males) were analyzed from the type locality in 2012 (23 larvae from 29 July, 8 from 3 July, 5 from 20 July). Chromosome I was metacentric and accounted for about 41% of the total complement length, whereas chromosomes II and III were submetacentric, with 30% and 29%, respectively, of the total length. The homologues were tightly paired. The centromere bands were heavily stained, and in chromosomes I and II they were located in conspicuously expanded regions. The nucleolar organizer was in the base of IIIL.
The short arm of chromosome I ( Fig. 5 View FIGURE 5 ) had the fine bands (section 1) characteristic of species in the genus Simulium , and was 2 fixed inversions removed from the sequence of S. muiscorum . The long arm of chromosome I (Fig. 6) included the ‘marker’ (section 25) and the Z marker (section 27), possibly homologous with the markers of the same names in the subgenus Simulium ( Rothfels et al. 1978) and the S. vernum group ( Hunter & Connolly 1986), respectively. IL had no fixed differences relative to S. muiscorum , but represented the sex arm (Fig. 7) of S. chimguazaense . IL-2, which was Y-linked in S. chimguazaense , was fixed in S. muiscorum . Chromosome IIS ( Fig. 8 View FIGURE 8 ) had the classic trapezoidal marker (section 53), ring of Balbiani (section 49), and bulges (section 42); its sequence was identical to that of S. muiscorum . The long arm of IIL ( Fig. 9 View FIGURE 9 ) included markers typical of the subgenus Simulium : the puffing band (section 57), grey band (section 60), jagged (section 63), and parabalbiani (section 67). It differed from the sequence of S. muiscorum by 2 overlapping fixed inversions. Chromosome IIIS ( Fig. 10 View FIGURE 10 ) included the typical blister (section 75), capsule (section 78), and 2 heavy band markers (section 75); its sequence was identical to that of S. muiscorum . The long arm of chromosome III ( Fig. 11 View FIGURE 11 ) had the basal marker, possibly homologous with the marker of the same name in the subgenus Boreosimulium ( Golini & Rothfels 1984) . IIIL was the most rearranged arm with respect to S. muiscorum ( Fig. 12 View FIGURE 12 ), differing by 3 overlapping fixed inversions. The base of IIIL represented the sex-differential region of S. muiscorum . We do not know which of the 2 species has the more central band sequence among related species of Trichodagmia , and therefore, we refrain from numbering the IIIL sections.
Simulium chimguazaense had two X-linked inversions (IL-1, IL-3) and one Y-linked inversion (IL-2) in 5 zygotic combinations (Figs. 6, 7); they assorted randomly in the combined sample from 29 June and 3 July ( Table 3 View TABLE 3 ). A sample from 20 July, possibly representing a different generation or cohort had 4 X1 Y 1 males and 1 X2 Y 1 male. At the type locality (29 June 2012), we found 1 female heterozygous for IL-1/IL-2 and 1 male homozygous for IL-1; neither is included in Table 3 View TABLE 3 .
X1 = IL-1; frequency = 0.558. X2 = IL-3; frequency = 0.442. Y1 = IL-2; frequency = 1.000.
In our sample of S. muiscorum , females (n = 10) were either homozygous (70%) or heterozygous (30%) for a heteroband in the extreme base of IIIL ( Fig. 12 View FIGURE 12 ), whereas all males (n = 7) were heterozygous for the heteroband. Two females also were heterozygous for IIIL-4 (= IIIL-1 of Moreno 1982). Our results regarding occurrence and frequencies of the heteroband and inversion agreed with those of Moreno (1982).
Other than sex-linked rearrangements, no polymorphisms were found in either S. chimguazaense or S. muiscorum .
FIGURE 6. Chromosome IL of Simulium chimguazaense (female larva), with X-linked inversion IL-1 present homozygously (X1X1). IL-1 is absent in S. muiscorum . Limits of X-linked inversion IL-3 and Y-linked inversion IL-2 are shown with brackets. IL-2 is fixed in S. muiscorum . C = centromere, ma = marker, Z = Z band.
FIGURE 7. Chromosome IL (distal ¾ only) of Simulium chimguazaense (female larva), showing the X1X2 sex-chromosome configuration.
DNA sequences. For COI, 510 of 710 sites were conserved, 184 were variable, and 121 were parsimony informative. COI sequences of the subgenus Trichodagmia had GC percentages of 34.3–35.4%, and the subgenus Simulium had 33.8–34.7%. For ITS2, 500 of 2642 sites were conserved, 173 were variable, and 73 were parsimony informative. The base composition for GC was 29.4–32.5% in Trichodagmia species and 20.4–39.7% in Simulium species. Genetic divergence between S. chimguazaense and S. muiscorum was 12%, which exceeds the divergence values between most species of insects ( Hebert et al. 2003, Ruiz-Lopez et al. 2012). Interspecific divergence values of 2.8–19.5% have been reported for species of the subgenus Trichodagmia , with the largest values occurring between members of different species groups ( Hernández-Triana et al. 2012). The consensus trees for COI did not group all Trichodagmia species in the same clade, whereas the trees for ITS2 revealed two clades, one with species of the subgenus Trichodagmia and the other with species of the subgenus Simulium ( Fig. 13 A–D). Both COI and ITS2 grouped S. chimguazaense with S. muiscorum .
Etymology. The species name “ chimguazaense ” is derived from the type locality in Chingaza, which (as “Chim-gua-za” or “Chim-wa-za”) in the extinct Muisca language means “God’s mountain of the night.” Types. Holotype: Male (pinned) with pupal exuviae in microvial of glycerin. Type locality: COLOMBIA, Department of Cundinamarca, Chingaza Natural National Park (PNN Chingaza), Diamante Sector, Chuza River (4°38'37.8"N 73°44'47.6"W), 3101 m asl, 29 June 2012, collected by A.S. Diaz and I.A. Lotta. Paratypes: 1 male and 2 females (pinned) with pupal exuviae in microvials of glycerin, 2 pupae and 2 larvae in 80% ethanol. Same data as for holotype.
Additional material examined. Pupae in 80% ethanol: 7 from same location as holotype, 3 July 2012, A.S. Díaz & F.A. Colorado; 26 from Diamante Sector, Diamante stream, 9–10 October 2011, A.S. Díaz & I.A. Lotta; 9 from Palacio Sector, 04°42'2.6"N 73°50'39.0"W, 5 April 2012, A.S. Díaz, I.A. Lotta & F.A. Colorado; 4 from Monterredondo Sector, Monterredondo stream, 04°37'41.1"N 73°43'27.4"W, 9 October 2009, A.S. Díaz & I.A. Lotta. Larvae in 80% ethanol: 48 from Diamante Sector, Diamante stream, 3 July 2012, A.S. Díaz & I.A. Lotta; 4 from Monterredondo Sector, Monterredondo stream, 9 October 2009, A.S. Díaz & I.A. Lotta; 11 from Palacio Sector, Piedras Blancas, 3 April 2012, F.A. Colorado & I.A. Lotta.
Bionomics. Simulium chimguazaense colonized rocky streams and rivers 2975–3100 m asl. Larvae were attached to rocks and debris in swift streams 3–12 m wide, with temperatures of 8.5–9.5 °C, pH of 6.5–6.6, and dissolved oxygen of 7.81–8.21 mg /l. Simulium chimguazaense was collected with Gigantodax chilensis (Philippi) , G. paramorum Wygodzinsky & Coscarón , S. cormonsi Wygodzinsky , S. furcillatum Wygodzinsky & Coscarón , S. muiscorum , and S. pautense Coscarón & Takaoka. One female and two male larvae of S. chimguazaense from the type locality were infected with unidentified mermithid nematodes.
A 100 S. (S.) verecundum 89 S. (S.) verecundum S. (S.) reptans
100 S. (S.) reptans
S. (T.) nigrimanum 100 S. (T.) nigrimanum S. (T.) chimguazaense n. sp. 100 S. (T.) muiscorum
0.07 0.06 0.05 0.04 0.03 0.02 0.01 0.00 B 99 S. (T.) guianense
S. (T.) chimguazaense n. sp. 91 S. (T.) muiscorum
0.12 0.10 0.08 0.06 0.04 0.02 0.00 C
67 S. (S.) verecundum S. (S.) reptans
100 S. (S.) reptans
S. (T.) nigrimanum 100 S. (T.) nigrimanum S. (T.) chimguazaense n. sp. 99 S. (T.) muiscorum
35 30 25 20 15 10 5 0
D
99 S. (T.) guianense S. (T.) guianense S. (T.) chimguazaense n. sp. 93 S. (T.) muiscorum S. (S.) tuberosum 100 S. (S.) tuberosum
25 20 15 10 5 0 Comparisons with related species. The hypogynial valves of the female of S. chimguazaense are most similar to those of S. sumapazense , based on the illustration by Muñoz de Hoyos (1996). The valves are thinner and more digitiform than the subtriangular valves of S. muiscorum . In S. chimguazaense the central, black stripe of the scutum is thicker than in S. muiscorum . The scutum of a pharate female of S. sumapazense was described as dark brown, evidently without a pattern (Muñoz de Hoyos 1996); well-tanned, dried material is needed to confirm the putative absence of a pattern.
No reliable characters are available to distinguish the males of S. chimguazaense and S. sumapazense . The male of S. chimguazaense can be distinguished from that of S. muiscorum by the dimensions of the ventral plate (in ventral view): about 2.0, versus 2.4, times wider than long, respectively. As in other high-altitude species of the Andes ( Wygodzinsky & Coscarón 1979, Mantilla et al. 2013), the hair on the postocciput, antennal scape, and palpomeres II and III of the male of S. chimguazaense is dark, dense, and long, compared with the condition in related species at lower elevations.
The pupa of S. chimguazaense is similar to that of S. sumapazense and S. muiscorum . The 5 known pupae of S. sumapazense have 12 filaments (Muñoz de Hoyos 1996), and the number of pupal filaments in S. muiscorum is consistently 12, whereas the 44 examined pupae of S. chimguazaense have 10 (53.4% of gills), 11 (28.4%), or 12 (18.2%) filaments. The gill filament formulas and branching patterns of S. chimguazaense and S. sumapazense are nearly the same; however, the compact gill with uniformly short, swollen filaments of S. chimguazaense contrasts with the loosely arranged, slender filaments, with two notably longer than the others, in S. sumapazense . The pupa of S. muiscorum has slender filaments arranged as (2 + 4) + (2 + 4), contrasting with the (3 or 4) + 2 + (4 to 6) of S. chimguazaense . The cuticle of the head and anterior thorax of S. chimguazaense and S. sumapazense is smooth, but is endowed with microtubercles in S. muiscorum and other 12-filamented members of the S. orbitale group. The anterior margin of the cephalic plate is typically crenulate in S. muiscorum , but not in S. chimguazaense or S. sumapazense .
The mature larva of S. chimguazaense can be distinguished most reliably from other species of the S. orbitale group, as revised by Hernández-Triana (2011), by the swollen filaments of the gill histoblast. It also differs from S. muiscorum and S. sumapazense in meristic characters, such as the number of hooks in the posterior circlet, although the extent of geographical and environmental variation in these characters is unknown.
Simulium chimguazaense is molecularly distinct from S. muiscorum , based on both the barcoding COI gene and ITS 2. Molecular material of S. sumapazense , however, was not available for analysis.
The polytene chromosomes of S. chimguazaense differ from those of S. muiscorum by eight fixed inversions (IS-1, IS-2, IIL-1, IIL-2, IIIL-1, IIIL-2, and IIIL-3; IL-2 is fixed in S. muiscorum and present only in males of S. chimguazaense ). Sex-linked rearrangements of the two species are found on nonhomologous chromosomes: I in S. chimguazaense and III in S. muiscorum . A mixed population in the Chuza River, without evidence of hybridization, indicated that S. chimguazaense and S. muiscorum are reproductively isolated. Although we did not have cytologically fixed material of S. sumapazense , we were able to make limited comparisons with the chromosomal analysis by Romero (2002), who examined 48 larvae from the Teusacá River; the maps of S. sumapazense , however, did not allow a band-by-band comparison. The nucleolar organizer of S. sumapazense , like that of S. muiscorum , is in the base of IIIS ( Fig. 10 View FIGURE 10 ), rather than the base of IIIL. Ectopic pairing of centromeres I and II or of all three centromeres occurred in the majority of individuals of S. sumapazense . We found no ectopic pairing in S. chimguazaense or S. muiscorum . The parabalbiani in S. sumapazense is putatively in the basal half of IIL (not discernible on the maps of Romero 2002), but in S. chimguazaense and S. muiscorum , which differ from one another in the parabalbiani region by two fixed inversions, it is in the distal one-third of the arm. Sex determination was not clarified for S. sumapazense , but possibly is based in IIIL, whereas for S. chimguazaense it is in IL (Figs. 6, 7) and for S. muiscorum , it is in IIIL near the centromere region ( Fig. 12 View FIGURE 12 ).
Overall, S. chimguazaense is structurally most similar to S. sumapazense , followed by S. muiscorum . These three species are found at the highest altitudes (> 3000 m) among the known species in the S. orbitale group. We conclude that morphology, chromosomes, and molecular sequences collectively and independently corroborate the species status of Simulium chimguazaense .

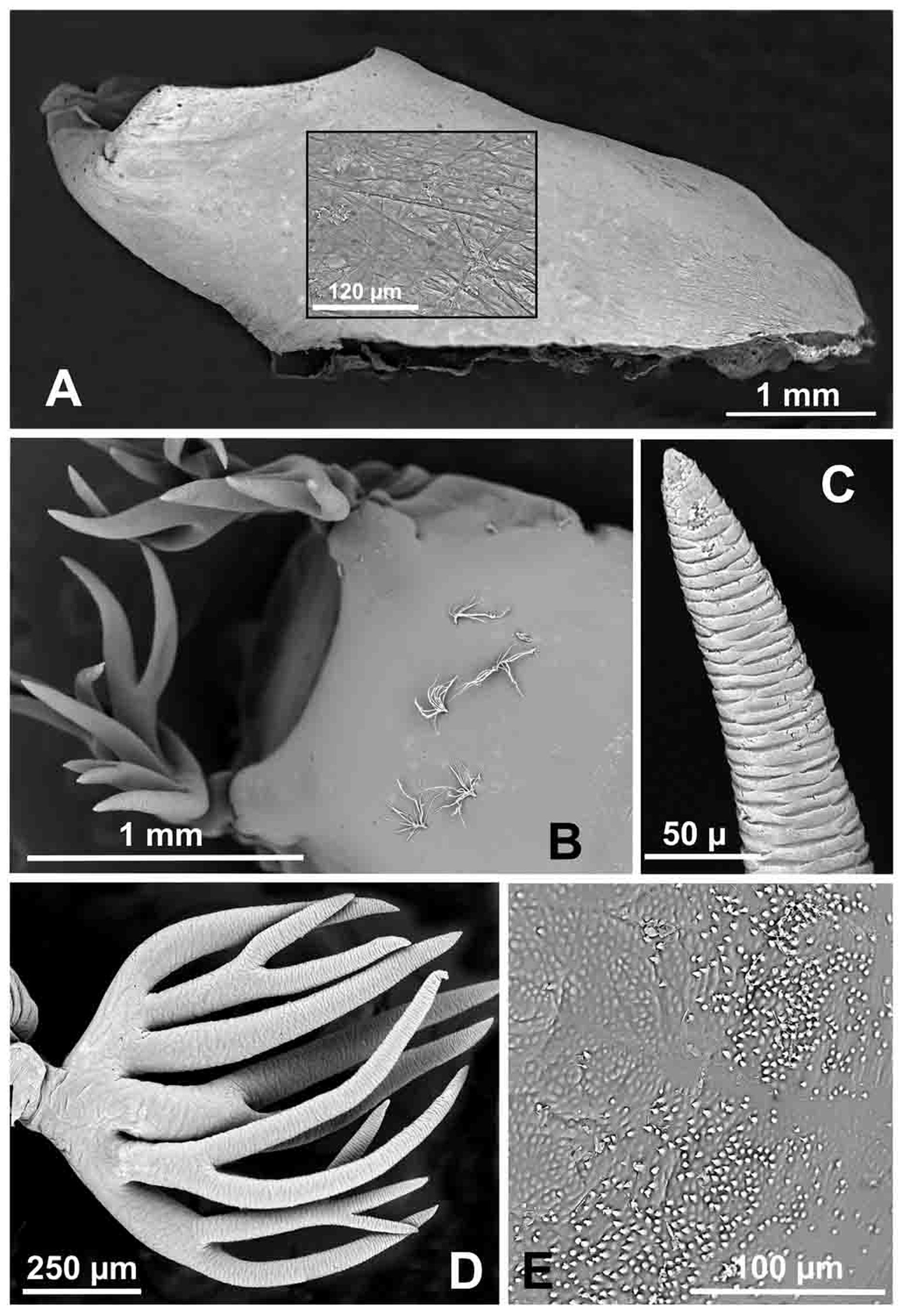


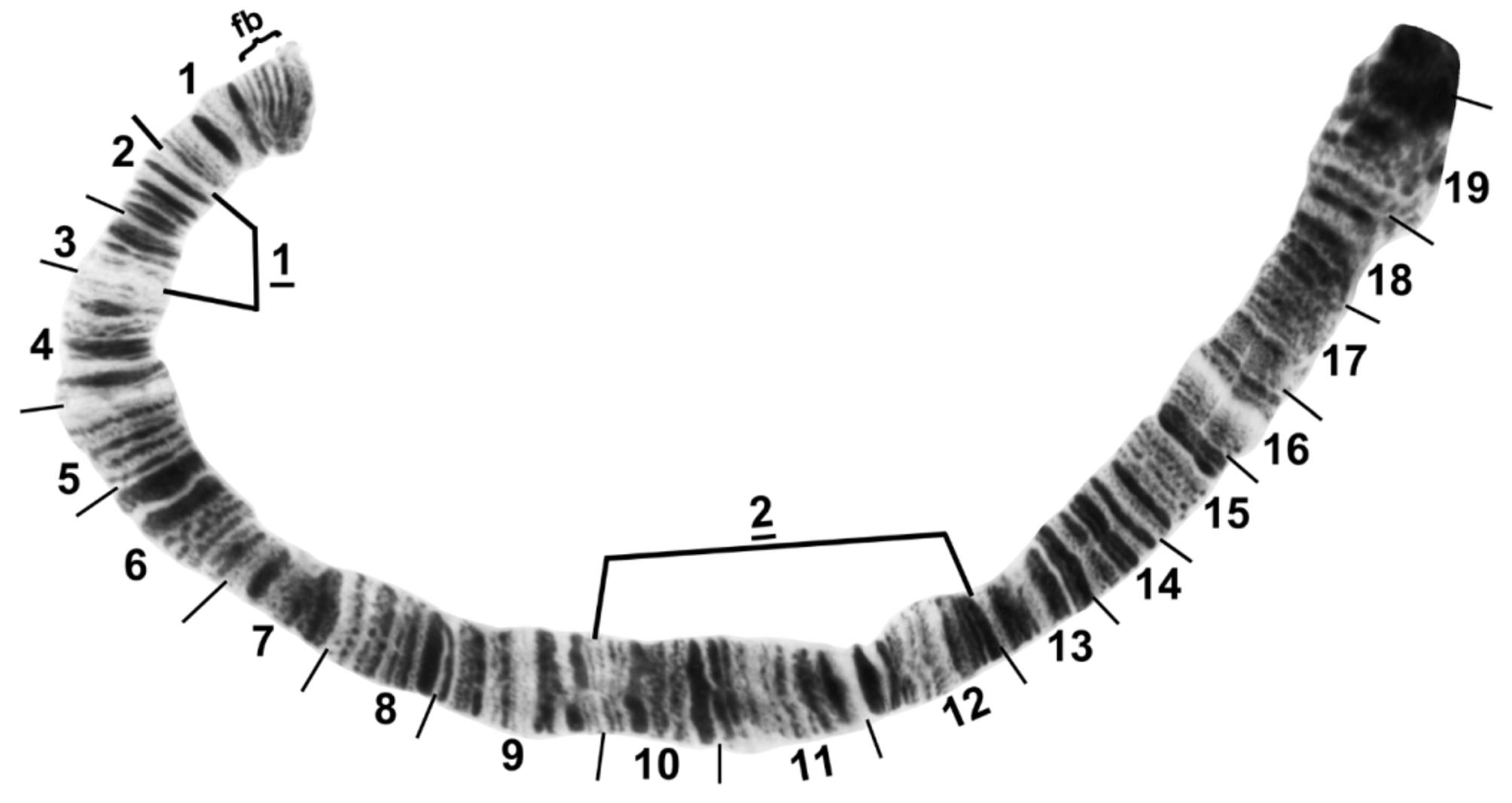
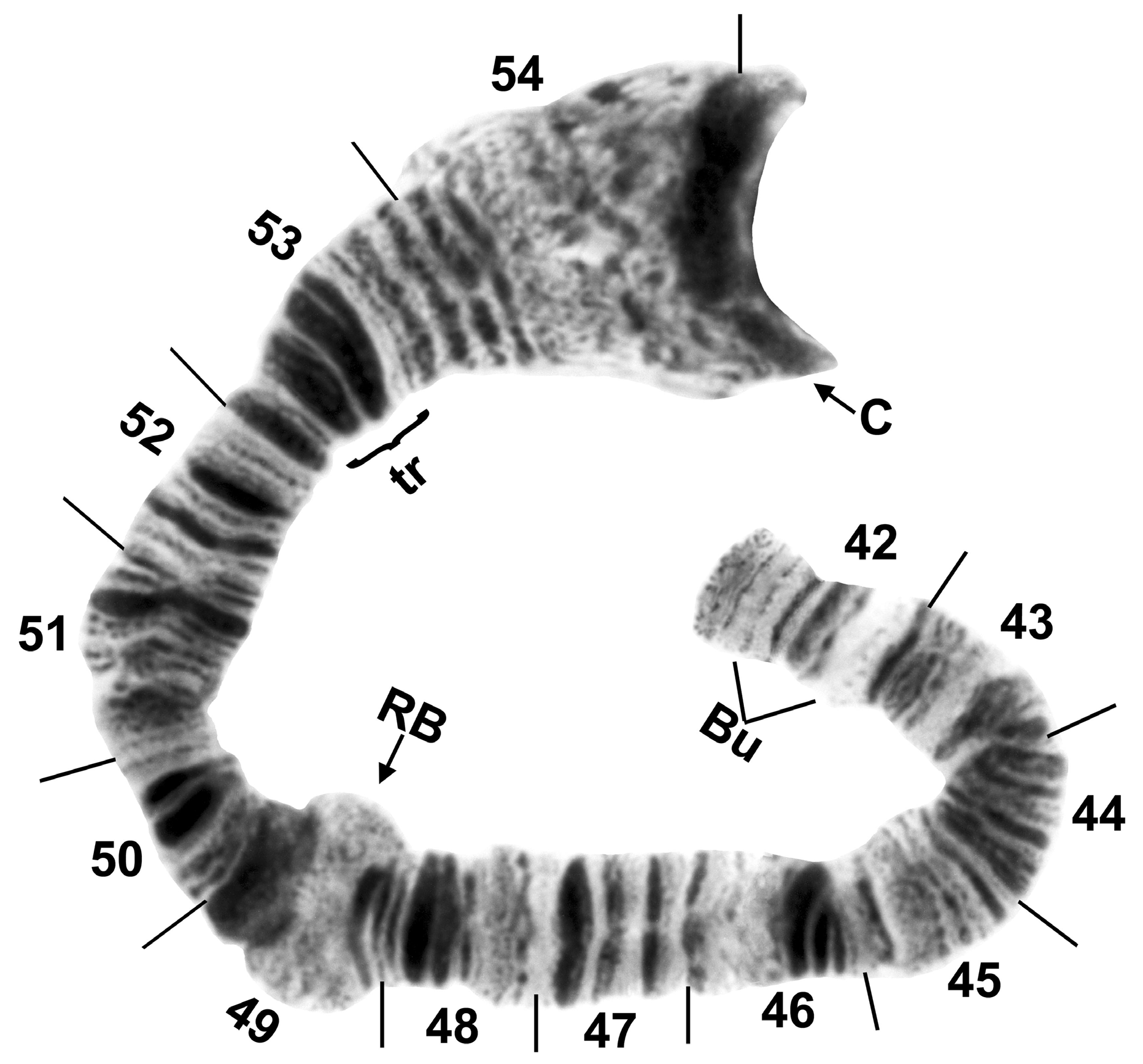
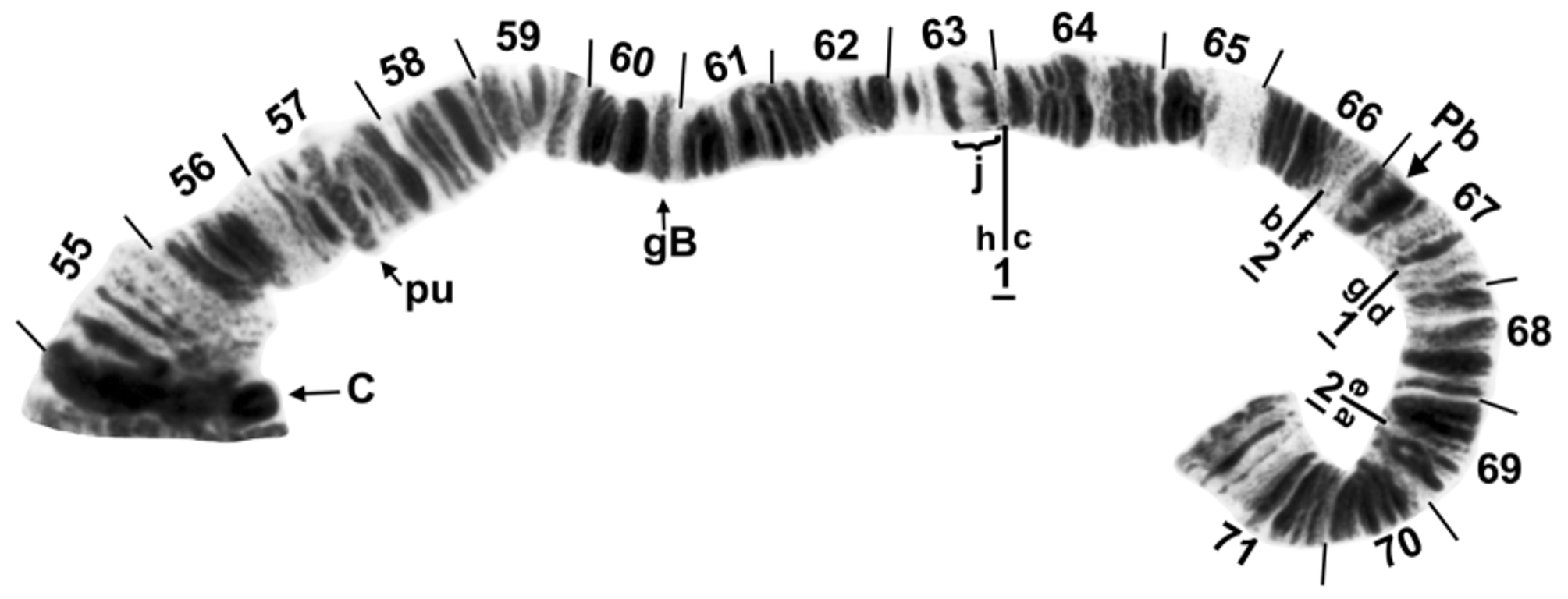


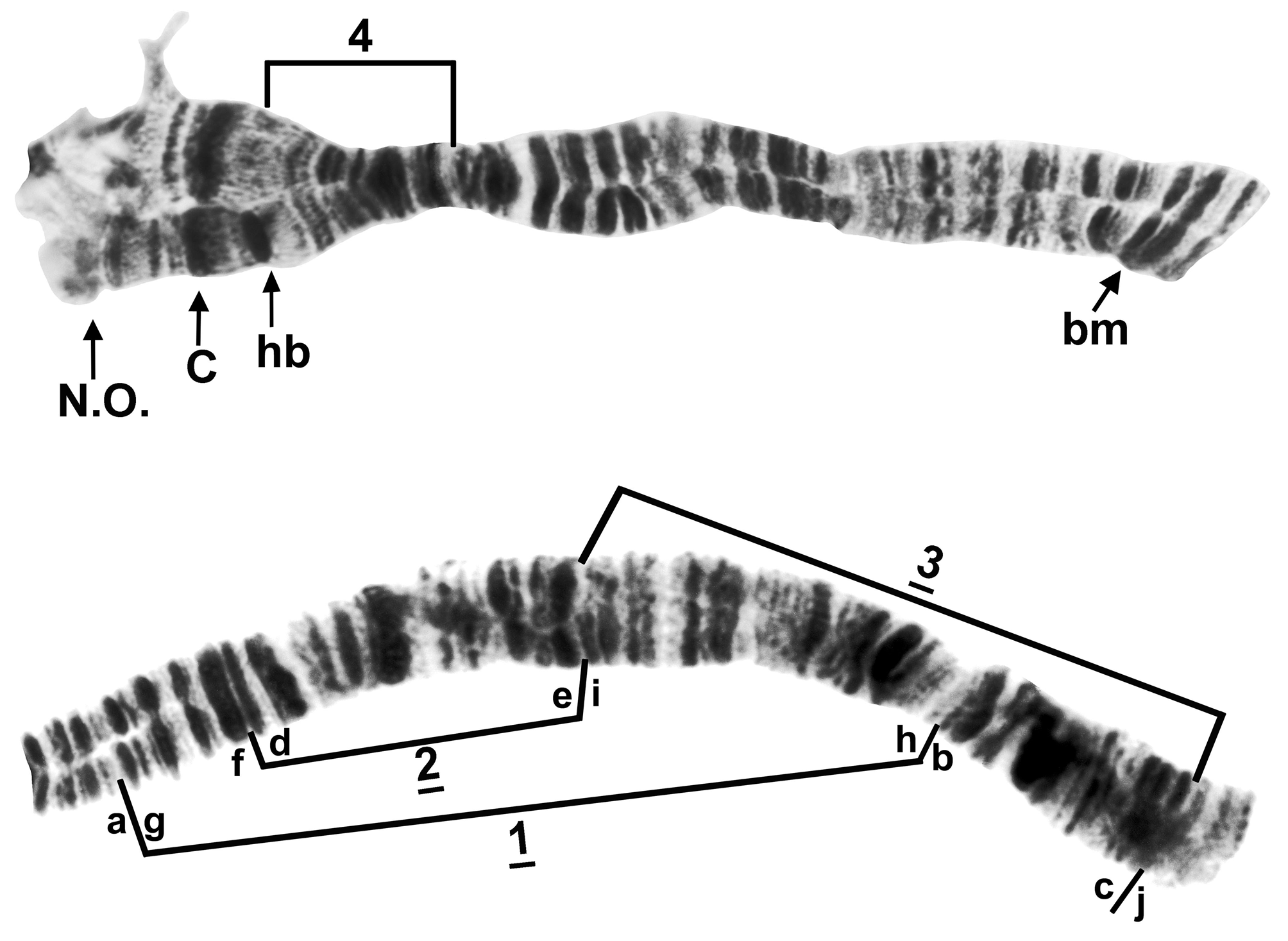
TABLE 3. G-test for random assortment of sex chromosomes in S. chimguazaense from the type locality, 29 June and 3 July 2012 (G = 1.2984, df = 4, P> 0.05).
| X1X1 | X1X2 | X2X2 | X1Y1 | X2Y1 |
|---|---|---|---|---|
| Observed 5 | 5 | 4 | 9 | 6 |
| Expected 4.36 | 6.91 | 2.73 | 8.37 | 6.63 |
| COI |
University of Coimbra Botany Department |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |