
Pseudochalcura alba Heraty & Heraty, 2009
|
publication ID |
https://doi.org/ 10.3897/zookeys.20.126 |
|
publication LSID |
lsid:zoobank.org:pub:32D10F5D-7DDA-426F-8CA9-E5538B7F6FD1 |
|
DOI |
https://doi.org/10.5281/zenodo.3791285 |
|
persistent identifier |
https://treatment.plazi.org/id/97405532-D55F-449C-9303-2F8BDFE1137E |
|
taxon LSID |
lsid:zoobank.org:act:97405532-D55F-449C-9303-2F8BDFE1137E |
|
treatment provided by |
Plazi |
|
scientific name |
Pseudochalcura alba Heraty & Heraty |
| status |
sp. nov. |
Pseudochalcura alba Heraty & Heraty , sp. n.
urn:lsid:zoobank.org:act:97405532-D55F-449C-9303-2F8BDFE1137E
Figs 1–9
Diagnosis. This is the only species of Pseudochalcura in which the male has flagellomeres 1–9 each with a long branch, the female has a long branch on flagellomeres 1–7, and both sexes have a 10-segmented flagellum. This species is placed in the condyla group of species based on the bare wing veins, sculptured (strigate) gastral sternite 1 in the female, absence of a metatibial spur, and ramose female antenna (to couplet 11 in Heraty 1986). No other member of this group has a 12-segmented female antenna or completely ramose male antenna. The closest species is considered to be P. prolata Heraty ( Argentina : Chaco Province), which has the basal 5–7 flagellomeres of the female antenna each with a stout branch, and a maximum of 8–11 antennal segments (male unknown). The basal branch of the female of P. prolata is stouter and 5.4 times as long as the basal length, as compared to 6.2 times in P. alba .
Male (Holotype). Length 2.35 mm. Black with faint metallic luster; scape, pedicel, mandible, petiole, and basal quarter of femora light brown; antennal flagellum and rest of legs white. Wings hyaline, stigma clear.
Head 1.79 times as broad as high (Fig. 4). Posterior ocellar line (POL) 2.8 times lateral ocellar line (LOL); POL 3.3 times ocellar ocular line (OOL). Frons and face irregularly costate (Fig. 4); clypeus and supraclypeal area smooth; genal bridge emarginate behind the mouthparts. Eyes separated by 2.2 times their height. Malar space 1.1 times height of eye, malar depression absent. Apical tooth of mandible long and overlapping opposing genal area. Labrum not observed. Antenna 12-segmented (Fig. 4); scape short and cylindrical, not reaching to median ocellus; all flagellomeres but the last with long branches ranging from 7.4 to 12.3 times as long as basal length, flagellomere branches alternating in origin from the base and slightly flattened; apical flagellomere unbranched and 3.6 times longer than broad; flagellomeres with dense short setae, and multiporous plate sensilla small and recessed into depressions of the antennal wall.
Figure Ι. Pseudochalcura alba sp. n., male holotype: habitus, lateral.
Mesosoma mostly with areolate sculpture, interstices broad with verrucose sculpture (Figs 2, 3, 5). Mesoscutum 2.0 times as broad as long; scutoscutellar sulcus transverse and deeply foveate; scutellar-axillar complex 1.4 times as long as maximum width of scutellum. Propodeal disc slightly rounded with broad alveolate sculpture medially and a few scattered setae dorsally, disc laterally relatively smooth; callus and metapleuron alveolate. Femoral groove deeply impressed. Proepisternum rugulose, becoming smooth apically. Mesocoxa weakly sculptured and lacking a lateral carina; metacoxa mostly smooth, laterally with small scattered pits. Legs stout (Figs 1, 2, 3), metafemur expanded medially, 3.5 times as long as broad; with short sparse appressed setae later-
Figures 2–9. Pseudochalcura alba sp. n.: 2– 5 male holotype: 2 lateral habitus 3 mesosoma, lateral 4 head, frontal 5 mesosoma, dorsal 6– 9 female paratype: 6 antenna, lateral 7 head, frontal 8 mesosoma, dorsal 9 habitus, lateral. Inset is magnification of mesoscutal sidelobe.
ally; metatibia with sparse semi-erect setae dorsally and dense appressed setae ventrally; metatibial spur absent. Forewing 1.9 times as long as broad; costal cell 0.31 times as long as wing, without setae; submarginal vein and basal area of wing bare; wing veins clear and difficult to discern beyond submarginal vein, stigma elongate oval and about 4 times as long as broad; disc of wing with microsetae ventrally. Hind wing broad and apically rounded, 3.4 times as long as broad.
Petiole 2.7 times as long as broad in lateral view, 1.6 times as long as metacoxa (Fig. 3); very slightly curved in profile; bare with fine granulate sculpture. Gaster globose, first tergite (Gt 1) 1.7 times as long as broad, smooth with few short setae dorsally; first gastral sternite smooth.
Female (Paratype; differences from male). Length 3.72 mm. Dark brown to black with faint metallic luster; antenna, mandible, petiole and basal two thirds of femora brown; rest of legs light brown.
Head 1.7 times as broad as high (Fig. 6). POL 2.5 times LOL; POL 2.8 times OOL. Frons and face finely costate (Fig. 7); clypeus smooth, supraclypeal area lightly costate. Eyes separated by 2.0 times their height. Malar space 0.77 times height of eye. Basal 7 flagellomeres with long branches ranging from 6.2 to 1.4 times as long as basal length (Fig. 6), flagellomere branches arranged linearly along flagellar axis, branches cylindrical; flagellomeres 8 and 9 lobate and with terminal segment forming a loose 3-segmented clava.
Mesosoma with areolate sculpture, interstices narrow and smooth (Figs 8, 9, and inset). Mesoscutum 2.1 times as broad as long; scutellar-axillar complex 1.1 times as long as maximum width of scutellum. Propodeal disc flat with broad alveolate sculpture medially, disc laterally areolate (Fig. 9). Metafemur 3.4 times as long as broad. Forewing 2.1 times as long as broad; costal cell 0.31 times as long as wing. Hind wing 3.4 times as long as broad.
Petiole 1.8 times as long as broad, 1.0 times as long as metacoxa; curved in profile, sculpture granulate with weak irregular carinae laterally. Gaster globose (Fig. 9), first tergite (Gt 1) 1.4 times as long as broad, smooth with few short setae dorsally; first gastral sternite longitudinally strigate with broad median keel. Hypopygium with two bristles. Ovipositor acicular.
Etymology. Named for the white antenna and light-colored petiole of the male.
Holotype: Argentina : Santiago del Estero Province, E of Lavalle , RN 64, 512m, 28°09’29”S 64°55’37”W, 10.III.2007, J. and J. Heraty, chaco vegetation H07-004, UCRC _ ENT 161499 View Materials ; deposited in UCR GoogleMaps . Paratype: Argentina : Catamarca, Punta de Balasto, 2100m, 25–26.I.1995, P. Fidalgo, UCRC _ENT 91260 (1 female, IFML).
Phylogenetic Results
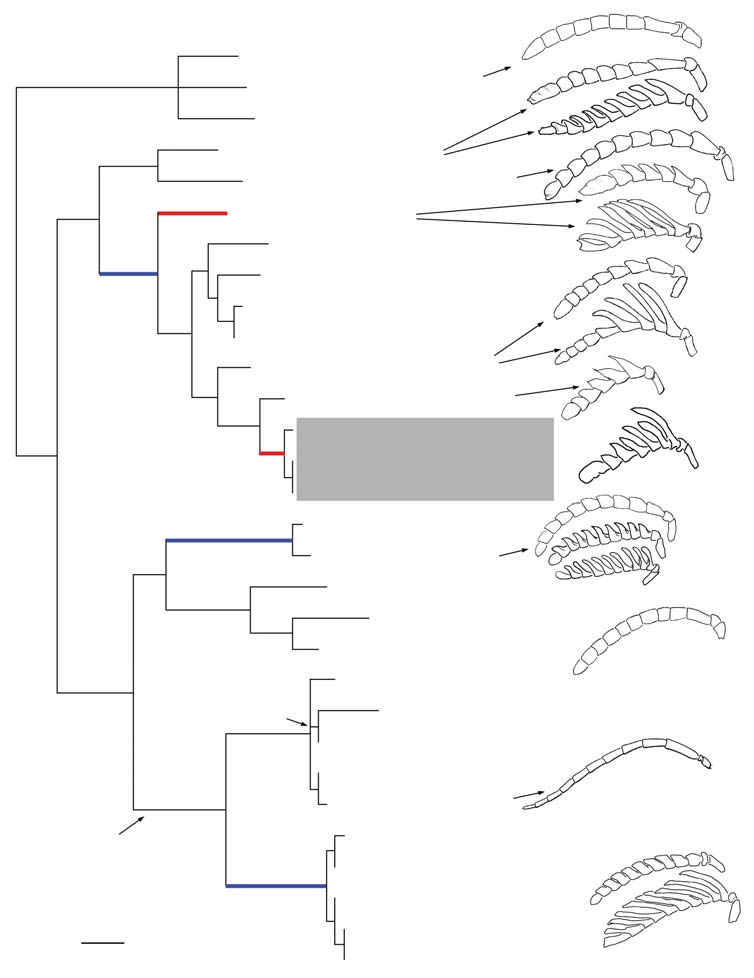
Only two genera of the Stilbula clade (sensu Heraty 2002) were not included in these analyses. Neostilbula ( Madagascar) is now considered as a member of the Eucharis clade in the Eucharitini (Heraty unpublished). Specimens of Striostilbula ( Australia) were not available for molecular analysis, although its placement in the Stilbula clade is based on weak morphological support and its inclusion is suspect ( Heraty 2002). Parsimony analysis of the 7 genera in the Stilbula clade resulted in a single tree (247 steps, r.i. 0.88; Fig. 23 View Figure 23 ). Successive approximations weighting generated the same tree, suggesting the data provide stable results ( Carpenter 1988). Bootstrap support was high for most nodes across the tree. The RAxML results provided nearly the same tree topology, but with Stilbuloida sister group to the rest of the Stilbula clade; however the RAxML bootstrap results supported a monophyletic Stilbuloida + Stilbula (100%; Fig. 23 View Figure 23 ). The RAxML results also placed P. gibbosa as sister group to the P. americana clade, but with almost no bootstrap support (52% as compared to 51% for the placement on the parsimony tree ( Fig. 23 View Figure 23 ). For Pseudochalcura , the results presented in Fig. 23 View Figure 23 are more concordant with the morphology-based phylogeny presented in Heraty (1986). Relationships within and among the other genera were the same in both results ( Fig. 23 View Figure 23 ).
Two groups occur within the Stilbula clade ( Fig. 23 View Figure 23 ): (A) Leurocharis (Australian) , Substilbula (Australian) , Rhipipalloidea (Indoaustralian) and Pseudochalcura (New World); weak parsimony bootstrap support (52%), but strong RAxML bootstrap support (99%); and (B) Stilbuloida (Australian) , Stilbula (Old World) , Obeza and Lophyrocera (both New World); strong parsimony bootstrap support (100%) but not monophyletic in RAxML. Genera in group B all share a bare callar region on the mesosoma, and usually have strong transverse carinae on the lower face. Sister group relationships between Rhipipalloidea + Pseudochalcura and Lophyrocera + Obeza have been proposed based on both morphological and molecular data ( Heraty 2002; Heraty et al. 2004). Rhipipalloidea and Pseudochalura have the postgenae or hypostomae forming a complete bridge posterior to the mandibles in association with a reduction of the maxillary complex ( Heraty 2002), and monophyly of Obeza and Lophyrocera share strong projections on the propodeum and callus ( Heraty 2002). Based on morphology, Leurocharis was treated as either a sister group to all of these genera when monophyletic, or exclud- ed when paraphyletic ( Heraty 2002). Subsequent analyses of an even larger molecular data set consistently put all of these Stilbula clade genera together in a monophyletic group (Heraty unpublished).
The new species, P. alba , is placed together with P. prolata Heraty ( Fig. 23 View Figure 23 , grey box). Pseudochalcura prolata was placed by Heraty (1986) with two other species, P. condylus Heraty and P. sculpturata Heraty , in the prolata species group based on six synapomorphies, including males having 7 basal rami on the antennal flagellomeres, the first gastral sternite striate, and lack of a metatibial spur, which are all features shared with P. alba . Pseudochalcura alba is the only male with rami on all flagellar segments, including an elongation of the terminal segment (Fig. 4). Females of P. alba , P. sculpturata , and P. prolata have at least the basal flagellomeres ramose (Figs 6, 14, 23g). Females of all other species of Pseudochalcura have either cylindrical or at most dorsally serrate antennae (Figs 10, 12, 16, 23e). Based on the lack of costal cell setae, P. pauca was treated as a potential sister group of the prolata group, but it has serrate, not ramose, antennae ( Fig. 23f View Figure 23 ); the male is unknown ( Heraty 1986). Pseudochalcura gibbosa , along with P. septuosa Heraty and P. atra Heraty , was proposed as the sister group of P. pauca + the prolata group ( Heraty 1986), which is consistent with the current results ( Fig. 23 View Figure 23 ). Males of the P. americana clade herein and P. gibbosa have only the basal 4–6 flagellomeres ramose (Figs 11, 13). Based on the phylogenetic hypothesis presented in Fig. 23 View Figure 23 , in males, a flagellum bearing 4–6 basal rami originating along the dorsal midline is considered plesiomorphic for the genus (Figs 11, 13), whereas a flagellum with 7–9 rami, with an alternating origin along the midline (Figs 4, 15, 17, 18) is apomorphic within Pseudochalcura .
Females of Rhipipalloidea from Australia have either simple cylindrical flagellomeres as in R. gruberi (Girault) (Fig. 19), dorsally serrate flagellomeres as in R. mira (Girault) (Fig. 20), or completely ramose flagellomeres as in several undescribed species from Sarawak (Fig. 21), Vietnam, and the Philippines. One undescribed species from Mudigere (western India) has dorsally serrate flagellomeres. Males of all species of Rhipipalloidea have long dorsal rami on flagellomeres 1–7 (Figs. 22, 23d). Female rami always originate along the midline, whereas male rami have alternating sites of origin. Thus the range of antennal morphology in these Australasian species parallels the diversity found in their New World sister group.
| UCRC |
University of California, Riverside |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |