
Sparassocynus
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3937.1.7 |
|
publication LSID |
lsid:zoobank.org:pub:4147764B-7FA0-475A-909C-E9FCA58E131C |
|
DOI |
https://doi.org/10.5281/zenodo.5613779 |
|
persistent identifier |
https://treatment.plazi.org/id/038987DA-536D-FFCC-FF5A-F89DFB7EFE6B |
|
treatment provided by |
Plazi |
|
scientific name |
Sparassocynus |
| status |
|
‘ Sparassocynus ’ heterotopicus Villarroel & Marshall, 1983
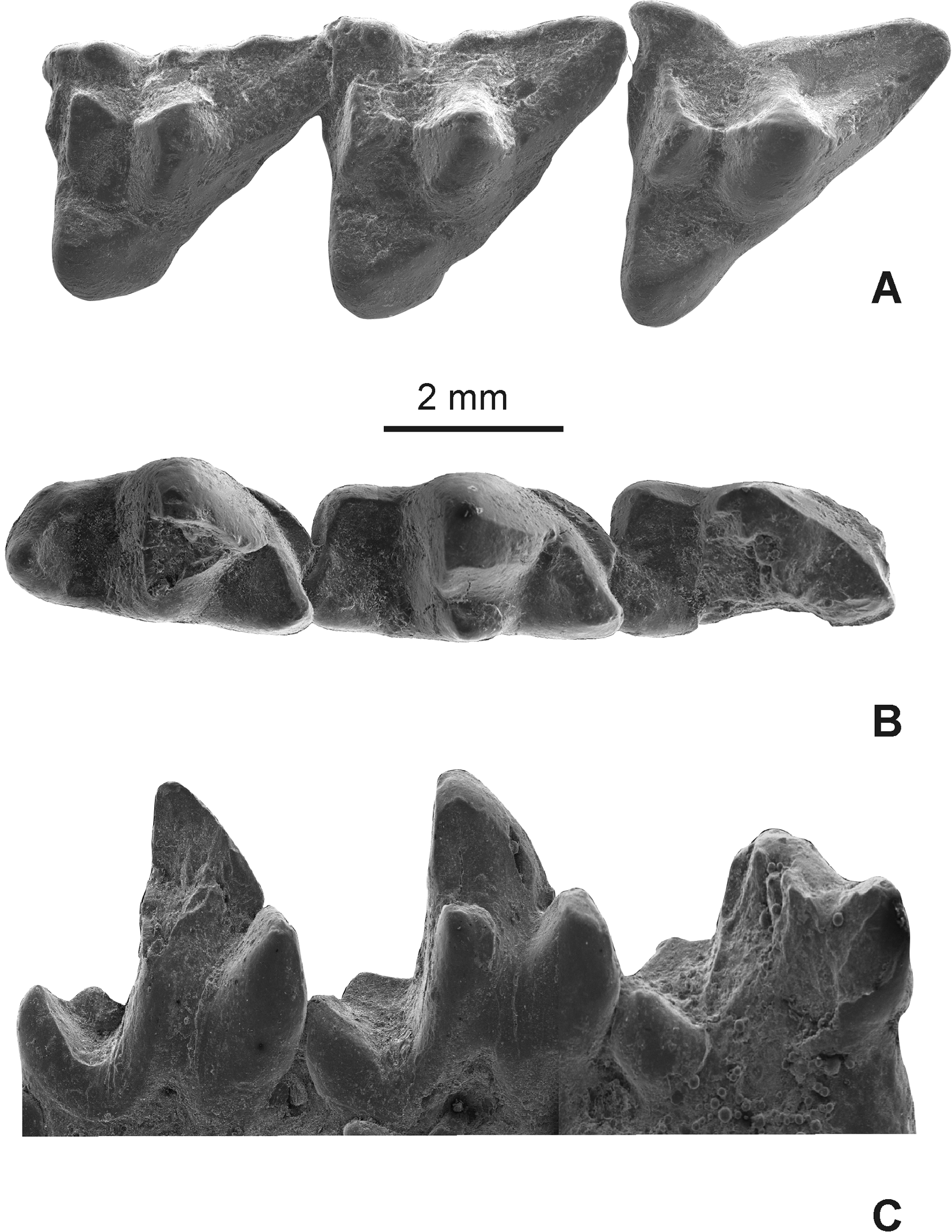
Fig. 4 View FIGURE 4 , Tables 1–2 View TABLE 1 View TABLE 2
Holotype. MNHN-BOL- 011896, fragment of a left maxilar with M1–3 ( Fig. 4 View FIGURE 4 A), fragment of a left mandible with m2–4 ( Fig. 4 View FIGURE 4 B–C), and an edentulous right mandible with roots of c–m4.
Locality and Horizon. Locality of Umala, Department of La Paz, Bolivia. Umala Formation (Montehermosan, early Pliocene sensu Marshall et al., 1979).
Comparative description. The general morphology of the dentition of S. heterotopicus shows characters common to diverse carnivorous didelphoids ( Goin & Pardiñas, 1996: 351; Forasiepi et al., 2009); among them are the upper molars with long metacristae and small protocones, and lower molars with small talonids and strong paracristids. Upper molars of MNHN-BOL- 011896 ( Fig. 4 View FIGURE 4 A) have a stylar shelf narrower than that of didelphids and as wide as in sparassocynids. The StA is absent on M1 and M3 and reduced on M2. On M1 the StB is the largest stylar cusp (in section it is half of the size of paracone), the StD is smaller than StB and subdivided into two cusps; finally StE is low and poorly developed. On M2 the StB is proportionally larger than on M1 and, as in the M1, StB is larger than StD; this latest is not subdivided and StE is missing. A reduced StB is present on M3. In contrast to the extant compared Didelphidae (Lutrolina, Didelphis , Monodelphis , Thylamys , and Caluromys ) and the extinct didelphids Thylatheridium , Thylophorops , and Hyperdidelphys , stylar cusps B and D are proportionally smaller. Compared to the Sparassocynidae , particularly Sparassocynus , the StB and StD are similarly developed, but MNHN-BOL- 011896 differs by having a subdivided StD on M1. Regarding relative sizes between StB and StD on M1–2, MNHN-BOL- 011896 is similar to the extinct didelphid Hyperdidelphis and sparassocynids in wich StB is larger than StD and differs from the extant compared didelphids, the extinct didelphid Thylophorops and Thylatheridium , by having a StB smaller than the StD. On M1 the preparacrista is short and points towards the parastylar corner whereas on M3 ends labially at the anterior base of the StB; on M2 the labial end of the preparacrista could not be evaluated because of wear. In the morphology of the preparacrista of M1, MNHN-BOL- 0 11896 differs from all compared taxa in which this crest joins the StB, excepting Didelphis , in which the preparacrista contacts the StA. On the other hand, the arrangement of the distal end of the preparacrista in the M3 of Umala specimen is similar to that of the M3 of Thylophorops , Hyperdidelphis parvula ( Goin & Pardiñas, 1996), and all compared extant didelphids (except Didelphis ); it also differs from the M3 of sparassocynids and Thylatheridium in which this crest joins the StB. The postmetacrista is long, sharp, and oblique as in carnivorous didelphids (e.g., Lutreolina ), but is slightly shorter than in sparassocynids (a long metacrista is a diagnostic character of the sparassocynids, Reig & Simpson, 1972, Goin, 1991). The metacone is large and the paracone is reduced as in other carnivorous forms ( Forasiepi et al., 2009); despite both cusps are closer to each other than in didelphids, they are not fused as in sparassocynids ( Reig & Simpson, 1972; Goin, 1991). The paracone is less reduced than the metacone compared to sparassocynids and didelphids. There is no evidence of para- and metaconule on M1–2, but on the M3 a small paraconule is present at the base of the paracone and the end of the preprotocrista. The protocone on M1–3 has a small occlusal surface in relation to the total molar occlusal surface; in this proportion, protocones are smaller than those of didelphids and larger than in sparassocynids. Furthermore protocones in S. heterotopicus are eccentric, similarly to those in sparassocynids and some didelphids (e.g., Thylophorops and Hyperdidelphys ). The ectoflexus is poorly developed in the M1–2 and strongest in the M3, as occurs, for example, in the sparassocynids and most didelphids ( Voss & Jansa, 2009).
The mandible of MNHN-BOL- 011896 is broken in front of the m2 and has lost most of the coronoid and angular processes, as well as a portion of the condylar process. The horizontal ramus is shallow (see Table 2 View TABLE 2 ) and bears a strong labial scar for the masseteric muscle on its posterolateral surface; the masseteric fossa appears to have been deep as in Lutreolina . The articular condyle is transversely elongated and the angular processes acute and strongly inflected as is common among didelphids ( Voss & Jansa, 2009). Lower dentition of MNHN-BOL- 0 11896 is partially preserved ( Fig. 4 View FIGURE 4 B–C). The metaconid, part of the protoconid of m2, and the metaconid of m4 are lost, so the morphology of the trigonid cusps (high and relative size) can be better described from those whole cusps of m3. Similarly to some didelphids (e.g., Thylatheridium ), the trigonid of the m3 is higher than the talonid, but not as tall as it is in sparassocynids. The paraconid is a robust cusp (as in m2 and m4), similar to the metaconid in height, but quite larger in section. The metaconid is a small cusp located close to the protoconid resembling sparassocynids and, in less extent, Lutreolina among didelphids. A strong paracristid is developed along the protoconid and paraconid cusps; it is not straight (e.g., Monodelphis , Thylamys , and Didelphis ) but forms an obtuse angle between the proximal (premetacristid) and distal (postparacristid) portions as occurs, for example, in sparassocynids, Thylatheridium and Lutreolina . The anterobasal cingulum of m2–4 is narrow and it extends posteriorly up to the anterior wall of the protoconid. The talonids of m2–4 are very shallow and have a smaller occlusal surface than trigonids; on m2–3 talonids are short and narrow like in the sparassocynids (Figs. 2, 4B and 5A; Table 2 View TABLE 2 ). In all molars the hypoconid projects slightly, not exceeding the labial wall of the protoconid. In the m2 both cusps are equally projected but in the m3 the hypoconid is placed more lingually than the protoconid, thus the talonid is narrower in the m3 than in the m2. In the m4 the hypoconid is a small cusp positioned behind the posterior wall of the protoconid. The cristid obliqua is slightly oblique on m2, and antero-posteriorly oriented on m3–4. The entoconid is a poorly differentiated cusp on m2–3 (better seen on m2), and absent on m4. The hypoconulid is smaller than the entoconid and positioned behind this cusp on m2–3. The talonid of m4 is relatively wider than in Sparassocynidae species and several didelphids such as Thylatheridium and Monodelphis . In contrast with these taxa, S. heterotopicus lacks a cingulum along the labial wall of the talonid bellow the hypoconid; in this regard, the talonid of m4 of S. heterotopicus is similar to that in Lutreolina and Didelphis .
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.