
Rhynchothorax Costa, 1861
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4567.3.1 |
|
publication LSID |
lsid:zoobank.org:pub:0AEFAF80-B001-4A18-88AC-5B6A189F6DCD |
|
DOI |
https://doi.org/10.5281/zenodo.5944898 |
|
persistent identifier |
https://treatment.plazi.org/id/03895C33-2907-4F1F-FF01-F975FCE9FE98 |
|
treatment provided by |
Plazi |
|
scientific name |
Rhynchothorax Costa, 1861 |
| status |
|
Genus Rhynchothorax Costa, 1861 View in CoL
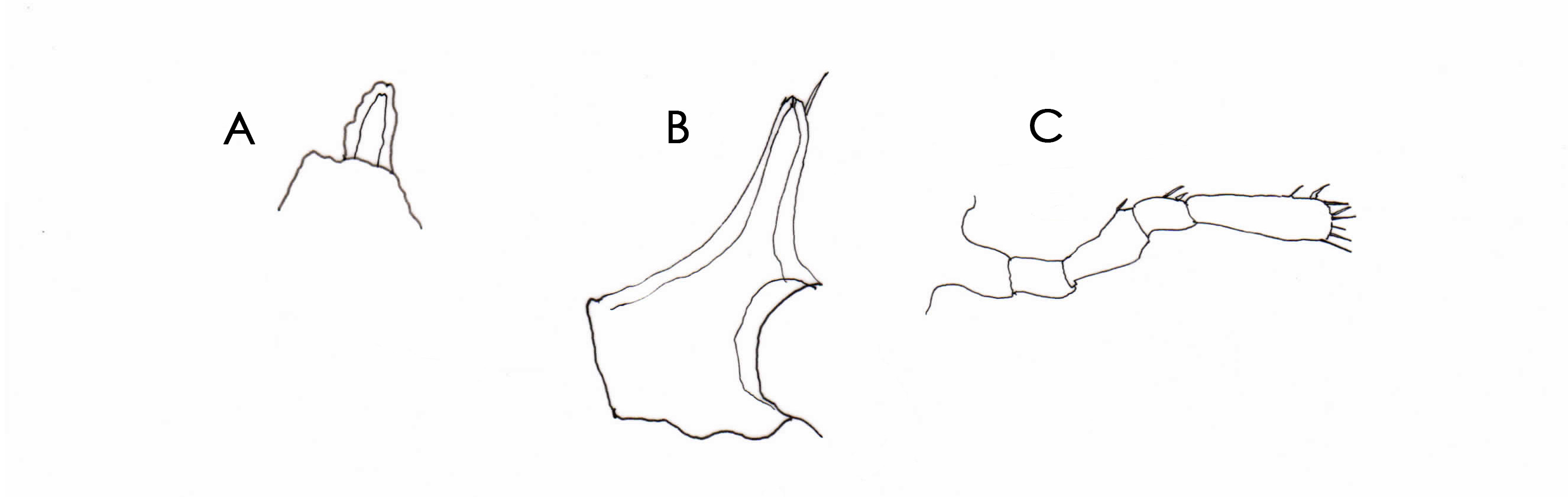
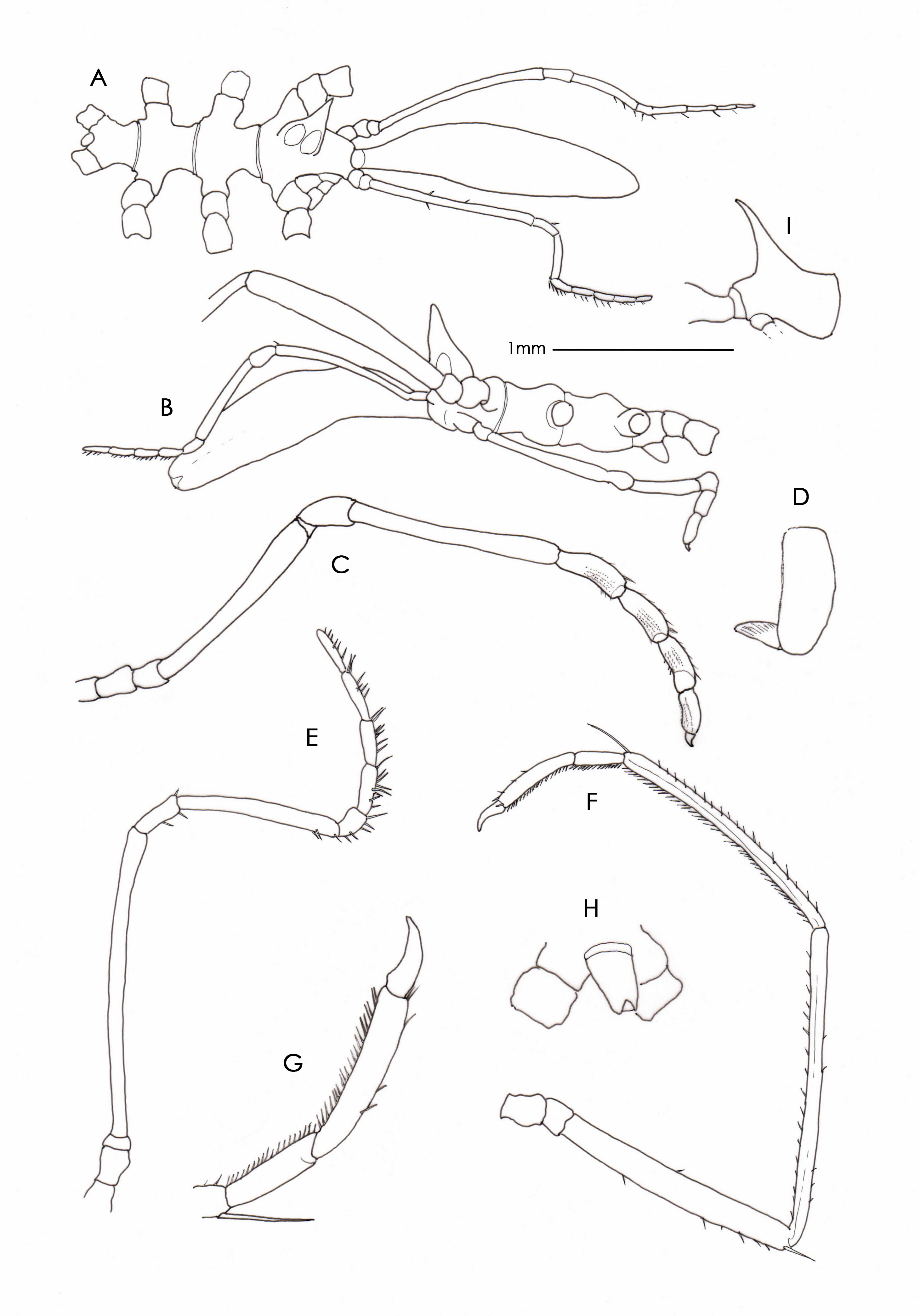
Remarks. Specimens of Rhynchothorax are mostly recorded interstitially. Nineteen species are currently recognized in the genus with four, five and six-segmented palps recorded. In this genus the palps are attached to lateral extensions of the cephalon. Confusion over the number of palp segments principally originates from historical uncertainty as to whether the lateral extension of the cephalon is segmented at its base. Child (1979) was the first to realize that the palps were attached to lateral extensions of the cephalon and that a number of authors had incorrectly figured a distinct suture line at the base of the lateral extension resulting in the lateral extension being counted as the first segment of the palp. This caused Child to reduce the number of palp segments by one in R. anophtalmus Arnaud, 1972 , R. barnardi Child & Hedgpeth, 1971 and R. philopsammum, Hedgpeth, 1951 and speculated that other species were similarly affected. Child (1995b) dismissed Hedgpeth’s (1951, plate 3c) illustration of a basal palp segment in R. philopsammum as an artifact and amended the number of palp segments in that species also from five to four. There is a further issue as to whether the lateral process is itself segmented due to a sometimes faint or incomplete segmentation line at its mid-length. This has resulted in instances where authors have interpreted that part of the lateral extension distal to the mid-constriction as the first or second segment of the palp. Child (1995b) proposed that this mid-constriction may represent the fusion of once distinct segments in the ancestral state. This constriction or suture appears to have first been illustrated in R. australis Hodgson, 1907 by Helfer (1938, Fig. 11 View FIGURE 11 ) as a dotted line in the mid-region of the cephalic lateral extension. Child (1995b) recorded five-segmented palps in R. australis and noted that a segmentation or suture line between the proximal short palp segments and the lateral cephalic extensions was very slight or missing entirely in many specimens and observed that the variable presence of this suture led to a miscount of palp segments in some individuals. Child (1995b, Figure 9B View FIGURE 9 ) illustrated a very distinct suture line and appears to have counted that part of the cephalic lateral extension distal to the suture line as the first of the five palp segments, and although he refers to ‘proximal short cylindrical segments’ he appears not to have counted a very short connecting segment that follows the lateral extension. On a similar note Child (1995b, fig. 10) illustrated four-segmented palps in R. percivali Clark, 1976 and showed no division of the lateral extension even though such a constriction appears to be quite evident in Clark’s SEM photograph ( Clark 1976, Fig. 1 View FIGURE 1 ). In this SWIR collection the suture line is missing or barely perceptible and any evidence of a constriction is mostly represented by a transverse band devoid of papillae. In any event there is no clear evidence of functional articulation. When assisted by strong back-lighting, the width of the cuticle wall lining the lateral extension is shown to be of continuous thickness throughout its length and with no evidence of thinning (or segmentation) at the point of the constriction. Clearly this current analysis must be provisional pending reexamination of some type specimens but based on the existing descriptions and material at hand I regard the cephalic lateral extension as not being segmented either proximally or at its mid-region. To add to the confusion there has also been uncertainty as to whether a tiny segment following the lateral extension represents a legitimate segment. There are many examples of confusion. Clark (1976, Figs 6 View FIGURE 6 , 15 View FIGURE 15 ) described and clearly illustrated a sixsegmented palp in R. percivali noting that the second short segment is easily overlooked. Child (1995b, Fig. 10 View FIGURE 10 ) on the other hand documented a five-segmented palp in the same species and noted that the second segment is longest. Child & Hedgpeth (1971) described R. barnardi with a six-segmented palp and clearly illustrated two basal segments. For reasons not fully discussed Child (1992b) later amended this number to four segments but noted a hint of a possible additional, extremely short, segment which he concluded to be connective integument rather than a segment itself. Nevertheless Child’s accompanying figure of the palp in lateral view does show more than a ‘hint’ of a well-defined segment ( Child 1992b, Fig.14A). This confusion is best demonstrated in R. australis Hodgson, 1907 where the five-segmented palp described by Hodgson (1907, Fig. V111, 3a & 1927, Fig. 17a View FIGURE 17 ) was amended to six by Calman (1915); the difference being that Calman (1915: Fig. 21) described and illustrated an additional “small but very distinct segment” between Hodgson’s first and second (longest) segment. Loman (1923) was the third author to address the number of palp segments in R. australis and in the absence of being able to find muscle structure on either side of Calman’s additional segment he dismissed it as being a true segment describing it as “a pseud-articulation (sic) (articulus spurious)”. Subsequently this segment has been clearly illustrated in R. articulatus Stock, 1968 by Clark (1976 Fig. 6 View FIGURE 6 ); in R. australis by Calman (1915, Fig. 21); in R. arenicolus Stock, 1989 ( Stock 1989 Figs 19 View FIGURE 19 ); in R. crenatus Child, 1982b ( Child 1982b, Fig. 167c) and R. percivali Clark, 1976 ( Clark 1976 Fig. 15 View FIGURE 15 ). Under high magnification it can be seen that this segment has a thickened cuticular wall and papillae on the surface which supports it being the first segment rather than connecting membrane. This segment is also wider than membrane connecting other palp segments and is itself bounded on both margins by narrow connecting membrane. The upward flexure of the second (longest) palp segment exposes the ventral surface of this tiny segment but at the same time partially or completely conceals its dorsal surface (see Child 1992b, Fig. 14A). The segment is therefore best seen in ventral or lateral view when the second (longest) segment is raised.
In summary I regard the cephalic lateral extension as not being segmented at either its base or throughout its length, and the tiny, often over-looked following segment that sits between the lateral extension and the longest segment, as the first segment of the palp. Based on these criteria and assuming there is uniformity in the structure of the basal palp segments in the genus, the palps of all species are five-segmented with three exceptions that need to be reviewed. These exceptions are R. crenatus Child, 1982b in which the palps are described as four-segmented and R. voxorinus Stock, 1966a and R. malaccensis Stock, 1968 in which the palps are described as six-segmented. It matters less as to whether my analysis is correct but rather that authors are cognizant of the issues involved and adopt a considered approach to future descriptions.
Until it is resolved as to which segment individual authors have regarded as being the ‘first’, the use of palp segments as a diagnostic character in the genus is problematical.





No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |