
Taimyria triassica NAUGOLNYKH et MOGUTCHEVA, 2022
|
publication ID |
https://doi.org/ 10.37520/fi.2022.018 |
|
DOI |
https://doi.org/10.5281/zenodo.7541197 |
|
persistent identifier |
https://treatment.plazi.org/id/03894B40-FFB1-FFB6-FF26-FCA8FEA78A52 |
|
treatment provided by |
Felipe |
|
scientific name |
Taimyria triassica NAUGOLNYKH et MOGUTCHEVA |
| status |
sp. nov. |
Taimyria triassica NAUGOLNYKH et MOGUTCHEVA sp. nov.
Text-figs 2–11 View Text-fig View Text-fig View Text-fig View Text-fig View Text-fig View Text-fig View Text-fig View Text-fig View Text-fig View Text-fig
H o l o t y p e. 4287/6, holotype figured here in Textfigs 2a, 3–10.
P l a n t F o s s i l N a m e s R e g i s t r y N u m b e r.
PFN003027.
R e p o s i t o r y. Monographic Department of the State Darwin Museum, Moscow, Russia .
During the study, the collection was kept in the Geological Institute of Russian Academy of Sciences (Moscow, Russia).
D e r i v a t i o n o m i n i s. From Triassic period.
Ty p e l o c a l i t y. The Tsvetkov Cape locality, Taimyr, Siberia, Russia; Lower Triassic, Induan.
D i a g n o s i s. Same as for the genus.
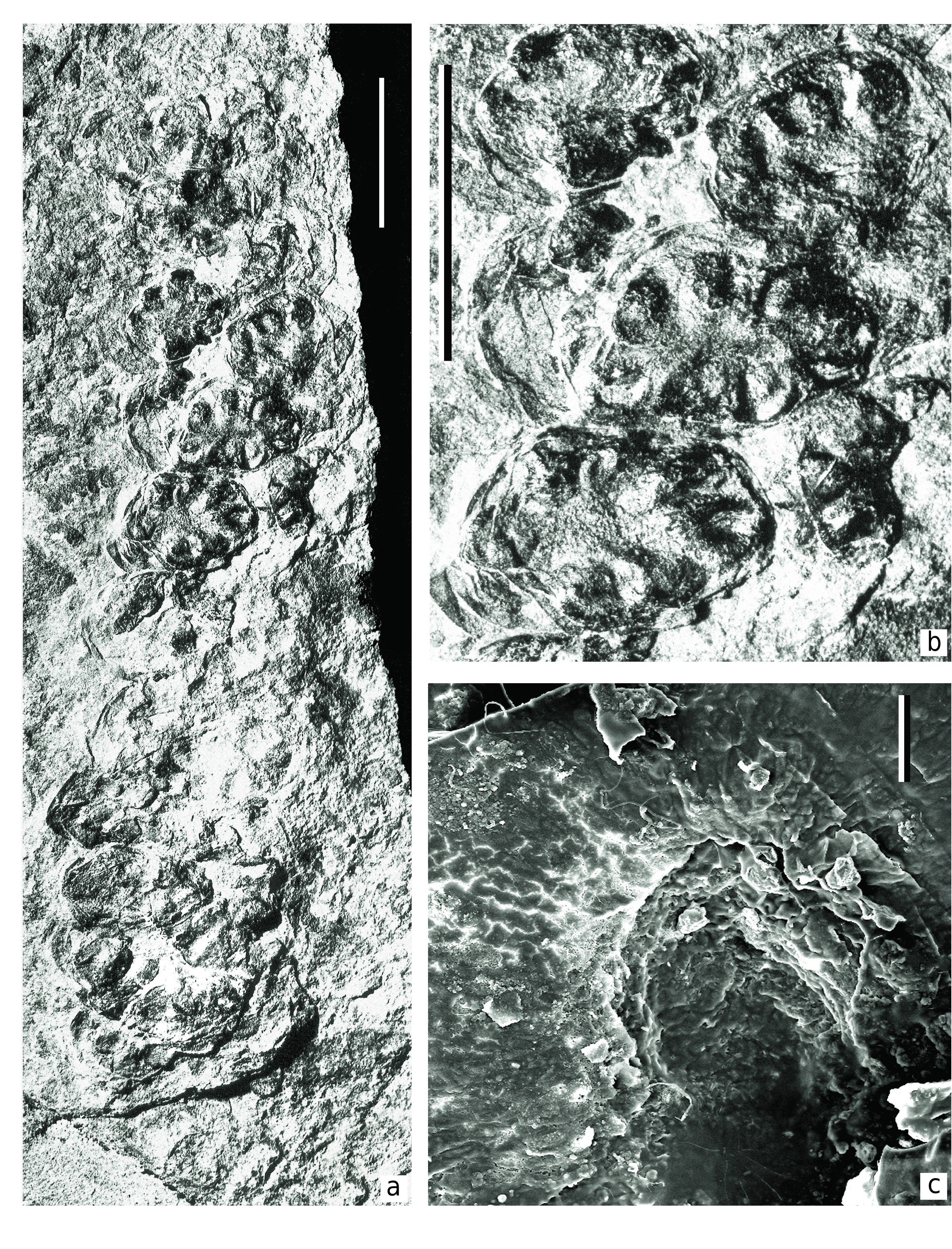
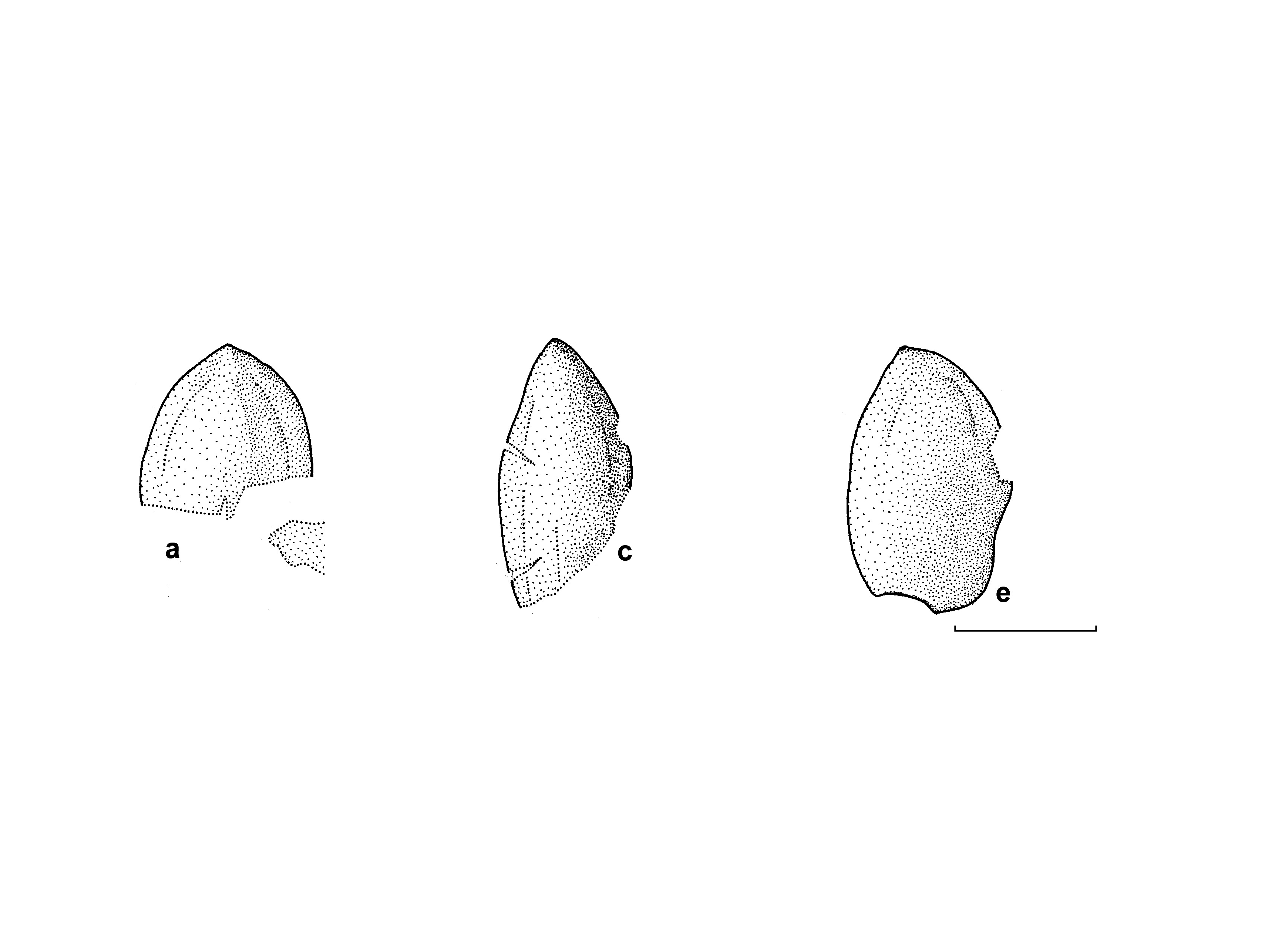
D e s c r i p t i o n. Macromorphology. The specimen studied is represented by four megastrobili (marked by arrows on Text-fig. 2 View Text-fig ) preserved on one bedding surface. There is a thick axis on one side of the specimen (Ax in Text-fig. 2 View Text-fig ). The four megastrobili are located on both lateral side of the Ax axis, with two on each side. Therefore, one can suppose that the megastrobili were arranged on a fertile branch in pinnate order, or forming a pinnate frond-like structure. Two megastrobili are preserved almost completely. Since the actual attachment of the megastrobili to the same branch (Ax) is not clearly visible, only one of the strobili is selected as a holotype ( Text-fig. 2a View Text-fig ). Two other strobili preserved on the other side of the branch are not complete and show their basal parts only.
The most well-preserved strobilus (holotype) is of more or less cylindrical shape ( Text-fig. 2a View Text-fig ). It is about 61 mm long and 15 mm wide. The base of the strobilus is not visible. Second strobilus, which is also preserved almost completely, is very similar to the holotype, but somewhat narrower, 72 mm long and 12 mm wide.
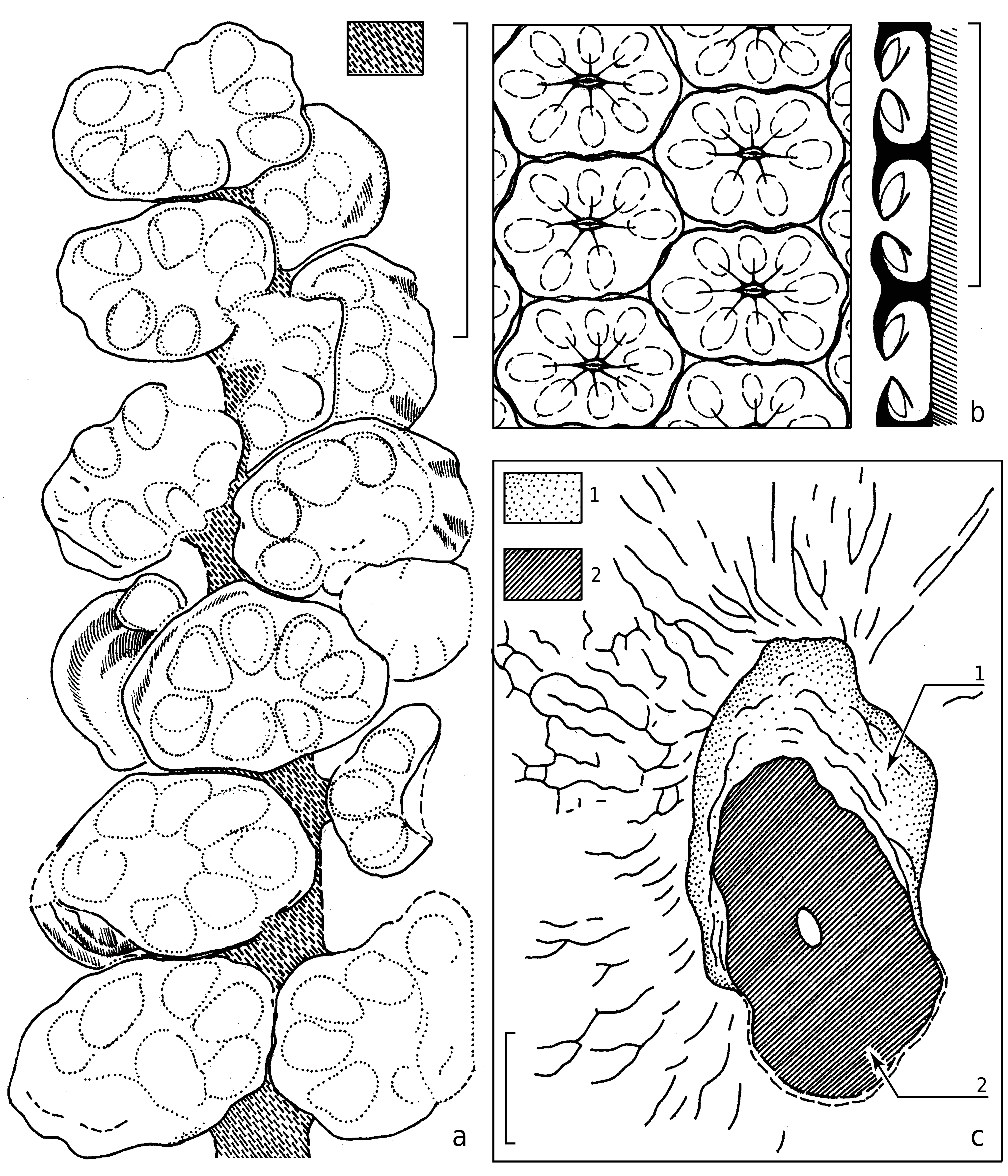
Each strobilus consists of densely arranged seed-bearing megasporangiate discs attached to the common axis in spiral order and forming well-pronounced parastichy ( Text-figs 3a, b View Text-fig , 4a, b View Text-fig ). Each observed parastichus on one side of the strobilus has three seed-bearing discs. Thus, one complete cycle should include six discs. The holotype has eleven parastichy. The second, narrower megastrobilus has fourteen parastichy. Thus, each strobilus has more than seventy seedbearing discs.
The possible fertile branch axis near the basal parts of the strobili is 7 mm wide and 80 mm long at visible part ( Text-fig. 2 View Text-fig , Ax). The surface of the axis is covered by very fine longitudinal ribs and furrows.
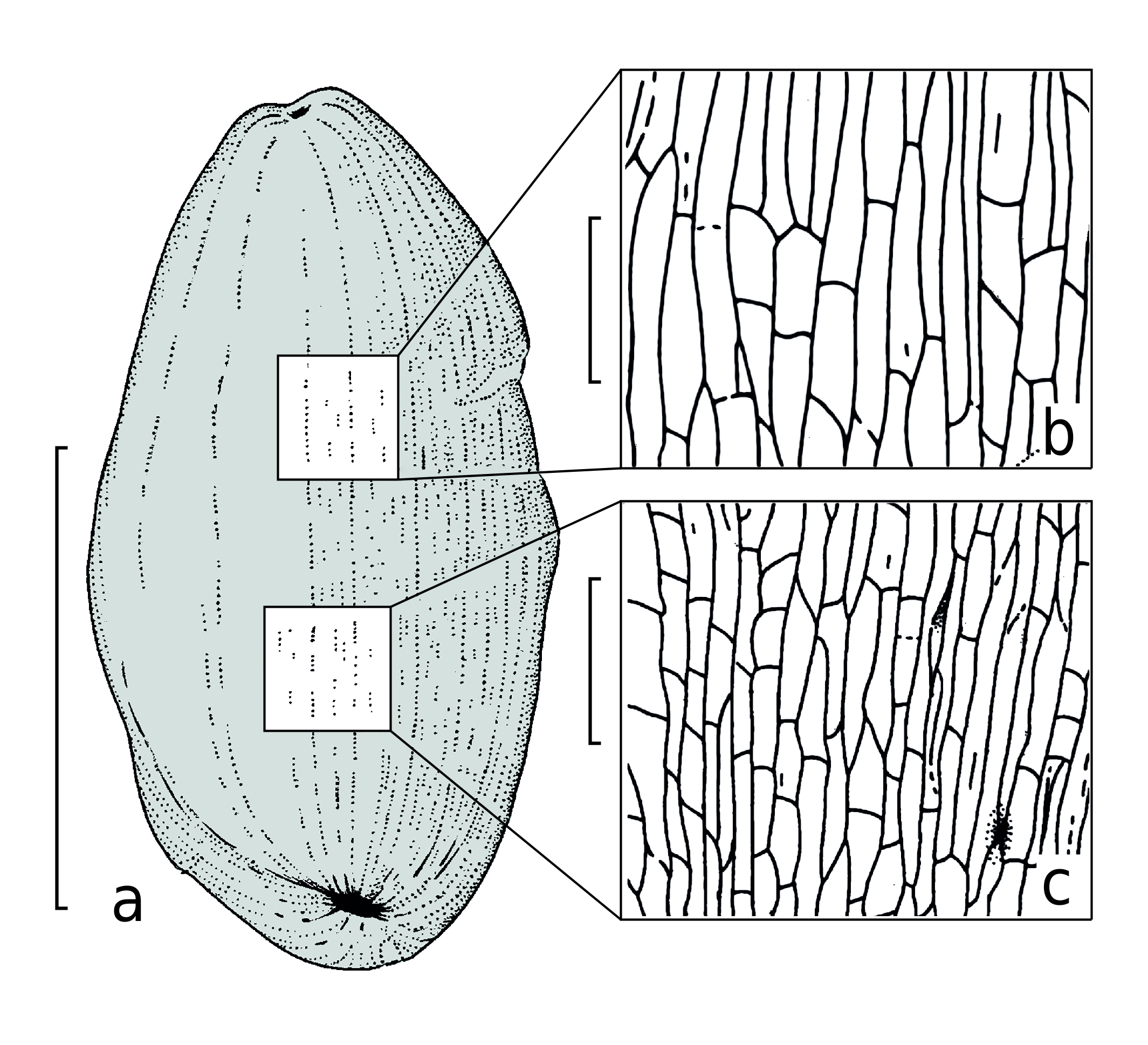
The seed-bearing discs are distinctly asymmetrical, ca. 5 × 9 mm in dimensions ( Text-figs 2a View Text-fig , 3a View Text-fig , 4a, b View Text-fig ). The long axes of the disc shields are oriented horizontally, and perpendicular to the main axis of the strobilus. This feature makes the present fossil different in comparison with other similar peltasperm female reproductive organs. The functionally upper or abaxial surface of the seed-bearing disc is smooth or slightly folded. Peripheries of the discs are entire-margined generally, but some discs bear six to eight unclear lobes about 1 × 2 mm in dimensions. Some discs demonstrate a flattened protective limb of about 2 mm wide at outer periphery of the lobes, making the disc semi-closed. This limb covers the seed base ( Text-fig. 4b View Text-fig ).
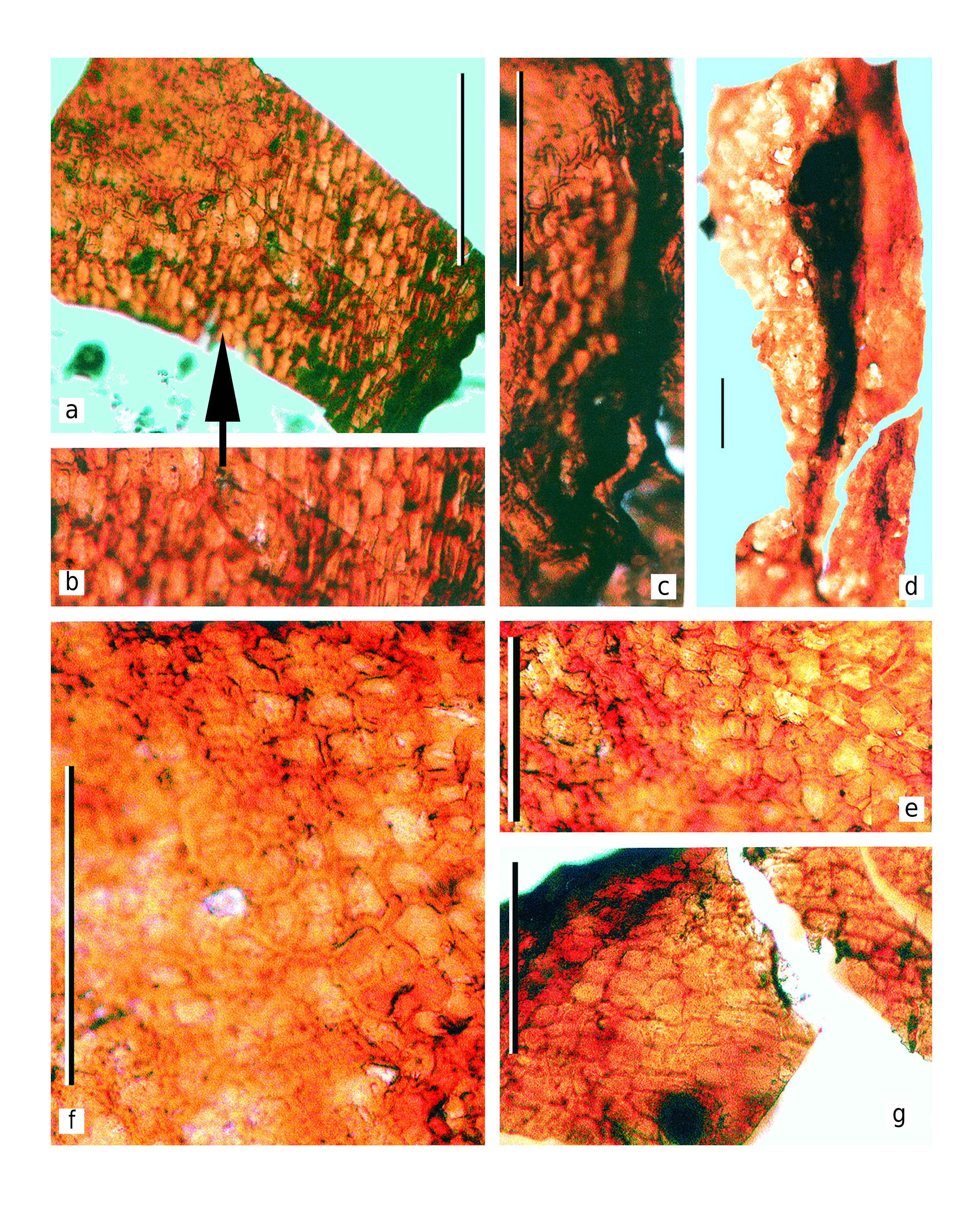
Cuticle preparations taken from the holotype show well-visible seed scars ( Text-figs 3c View Text-fig , 4c View Text-fig ). The seed scars are ovoid, 300 µm long and 200 µm wide on average. Size and shape of the seed scars well correspond to the size and shape of the chalazal base of the seeds. Judging from the material in hand, the seed scars were orientated along the radial directions of the seed-bearing disc.
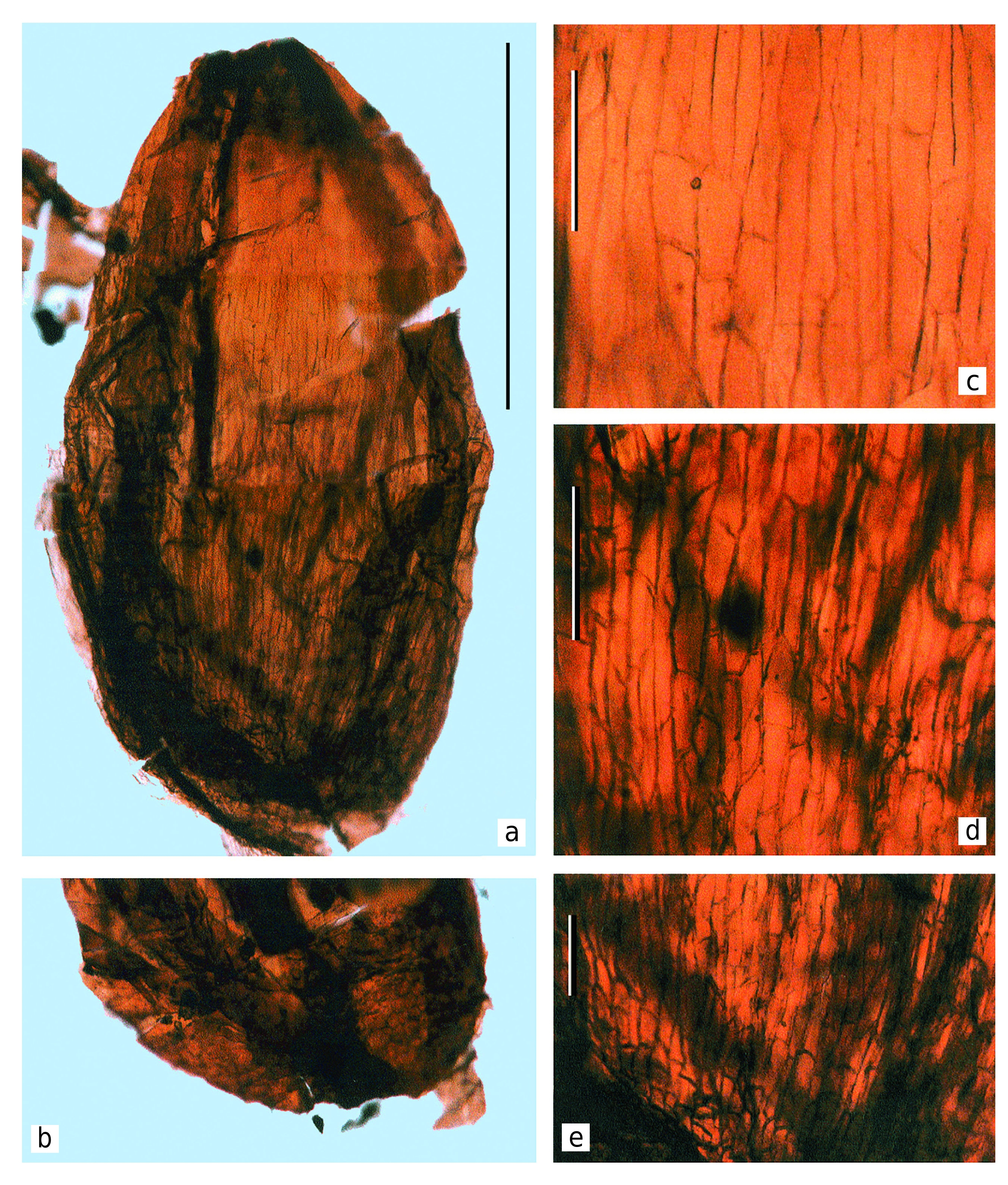
The seeds are disposed beneath the seed-bearing disc and directly attached to the adaxial surface of the discs ( Text-fig. 4b View Text-fig ). The seed cuticles were extracted during the maceration of the discs ( Text-figs 8–10 View Text-fig View Text-fig View Text-fig ).
The seeds are platyspermic, very small (according to the seed size ranking scale proposed by Artjushenko 1990). Seed are ovoid, 1.7–2.0 mm long and 1.0– 1.2 mm wide, widest at the middle or lower one-third ( Text-figs 8–10 View Text-fig View Text-fig View Text-fig ). The seeds are narrowed at the apical micropylar end. The seed peripheries are slightly flattened. The seed coat (spermoderm) is smooth, without distinct sculpture. The seed scarlet is about 300 µm in diameter, ellipsoidal to round, central in relation to the ventral side of the seed, somewhat impressed, surrounded by weakly developed ring-shaped uplifting ( Text-figs 8b View Text-fig , 9a View Text-fig ). The main axis of the seeds is straight to slightly curved. Since the seeds lack sarcotestal wings, we can conclude that the original plant had rather limited dispersal ability (authochorous or barochorous).
Very similar seeds, but considerably larger, were described as “ Cordaicarpus ” uralicus KH.R.DOMBR. from the lower Permian (Kungurian) deposits of the Cis-Urals ( Dombrovskaya 1976). It was suggested that these seeds were produced by the peltasperm plant with the fronds Permocallipteris retensoria (ZALESSKY) NAUGOLNYKH and the seed-bearing discs Peltaspermum sp. ( Naugolnykh and Kerp 1996, Naugolnykh 2014). Seeds of this type were found later in natural connection to the seed-bearing discs of Peltaspermum petaloides NAUGOLNYKH ( Naugolnykh 2016: 86, text-fig. 35A, B, D, pl. XXIX, 6).
Discovery of the present specimen, bearing seeds in natural connection to the seed-bearing discs similar to other representatives of Peltaspermales , supports the initial idea that these seed forms belong to the peltasperms ( Naugolnykh and Kerp 1996).
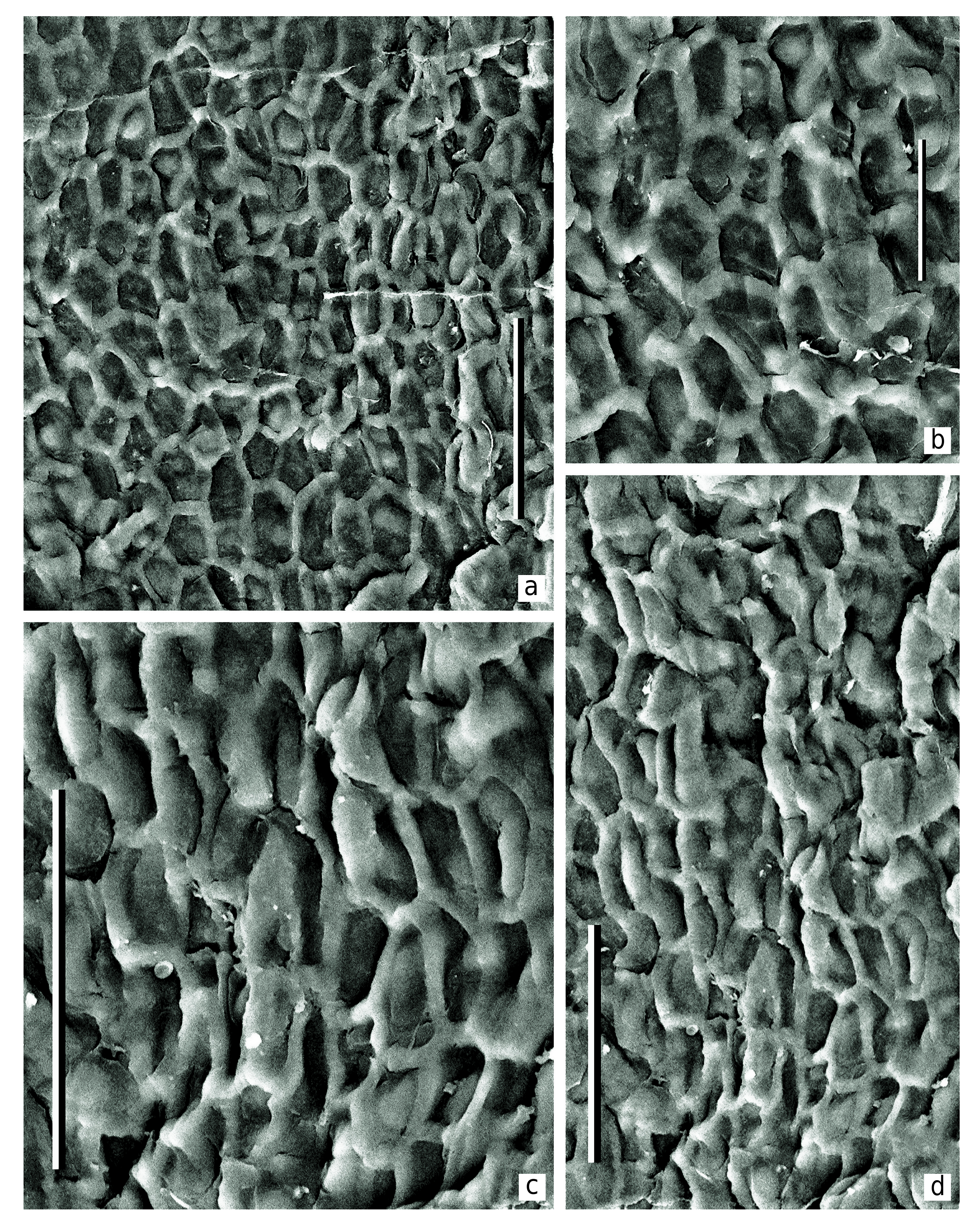
Epidermal-cuticular structure. The cuticles are relatively thick.The cell outlines are indistinctly pronounced.
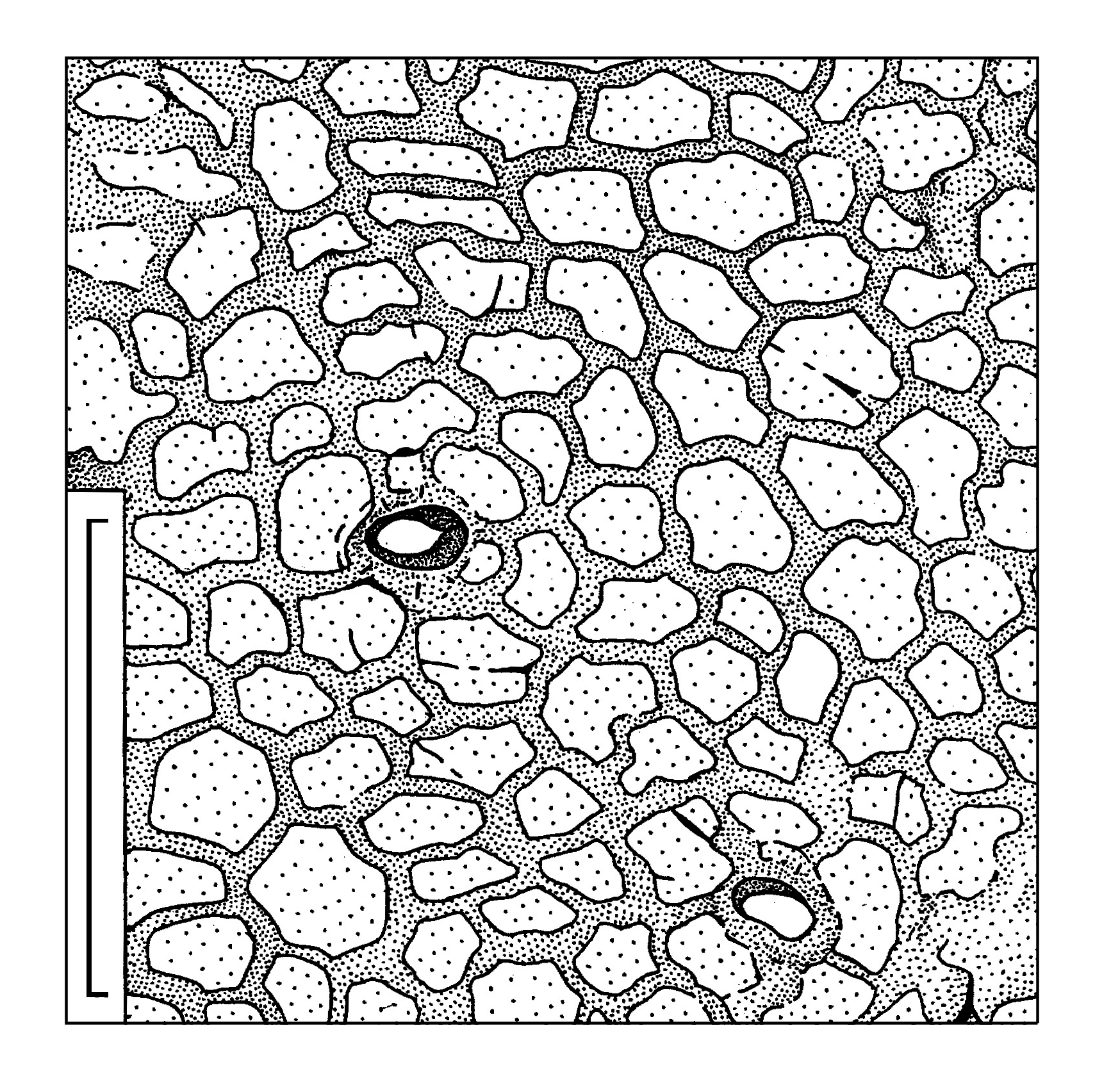
The most well-preserved cell structure is observed on the adaxial cuticles, especially around the seed scars, where the cells are clearly visible and well-expressed in the cuticle relief ( Text-figs 3c View Text-fig , 4c View Text-fig , 8–10 View Text-fig View Text-fig View Text-fig ).
The common epidermal cells are elongate, mostly of prolonged rectangular outlines, rarely more isometric. Average size of the cells is 40 × 70 µm. Radial cell walls are undulated to slightly curved. The straight cell walls are present as well. The periclinal cell walls are fairly smooth. The common epidermal cells form distinct rows radially disposed around the seed scars ( Text-figs 3c View Text-fig , 4c View Text-fig ). The cells become more isometric in the distance of 500–600 µm from the seed scar. Occasionally the papillae are joined into a common uplifting with weak extrusion ( Text-fig. 7 View Text-fig ). Similar cutinization of the stomata is known for some other representatives of the order Peltaspermales (e.g., Naugolnykh 2005: figs 12–14, 16, 17). In very rare cases, a crescent-shaped uplifting is developed instead the papillae. Common cells of the abaxial surface of the seed-bearing discs are of isometric outlines, round to polygonal, with average size 50 × 60 µm.

| R |
Departamento de Geologia, Universidad de Chile |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.