
Aliterella vladivostokensis Sh.R. Abdullin, A.Yu. Nikulin, V.B. Bagmet et V.Yu. Nikulin, 2021
|
publication ID |
https://doi.org/ 10.11646/phytotaxa.527.3.7 |
|
DOI |
https://doi.org/10.5281/zenodo.5753010 |
|
persistent identifier |
https://treatment.plazi.org/id/03884E68-FFA7-DD5F-3FC8-FD1BFDBF08F5 |
|
treatment provided by |
Plazi |
|
scientific name |
Aliterella vladivostokensis Sh.R. Abdullin, A.Yu. Nikulin, V.B. Bagmet et V.Yu. Nikulin |
| status |
sp. nov. |
Aliterella vladivostokensis Sh.R. Abdullin, A.Yu. Nikulin, V.B. Bagmet et V.Yu. Nikulin sp. nov. ( Fig. 1 View FIGURE 1 )
Description: —Cells solitary, more commonly irregular or in rounded colonies with many cells (up to 32–64 or more), usually aggregated irregularly, extended ( Fig. 1 A–C View FIGURE 1 ). Mucilage unstratified, colorless and firm, surrounding cells and colonies. Cells cylindrical 1.6–6.87 μm long, 1.17–5.85 μm wide, 1 to 1.75 × longer than wide (mean, 1.28 ×) ( Fig. 1 C–D View FIGURE 1 ). Cells easily squeezing from colonies with pressure ( Fig. 1 C View FIGURE 1 ). The chromatoplasm and centroplasm usually recognizable with light microscopy ( Fig. 1 C–D View FIGURE 1 ). Cell contents blue-green, slightly granulated, or sometimes homogeneous. Reproduction by simple binary cell division in three or more planes.
Holotype: —The dried biomass of authentic strain was deposited to the Herbarium of the Federal Scientific Center of East Asia Terrestrial Biodiversity, Russia (exsiccatum number VLA-CA-1212) as a holotype.
Type locality: — RUSSIA. Primorsky Territory, Vladivostok, 43°10’21.4” N, 131°56’10.9” E, collected by Shamil R. Abdullin on August 7, 2018. The authentic strain VCA-43 (Vl15-3) is available in the culture collection of the Laboratory of Botany, Federal Scientific Center of East Asia Terrestrial Biodiversity , Russia GoogleMaps .
Habitat: —This cyanobacteria occurred in aerophytic habitat (on the concrete fence).
Etymology: —The species epithet ‘ vladivostokensis ’ is derived from the type locality Vladivostok City.
Molecular phylogeny and sequence analyses: —Results of the BLAST searches showed that the sequence of the 16S rRNA gene and internal transcribed spacer (ITS) region (1800 bp) in our strain was highly similar to those in other species of Aliterella , several uncultured bacterial clones, and Synechocystis sp. PCC 7509 (> 94.5%). When only the ITS region was compared, the similarity to Aliterella spp. was just above 85%. Such relatively low similarity percentages suggested that we were likely dealing with a new species.
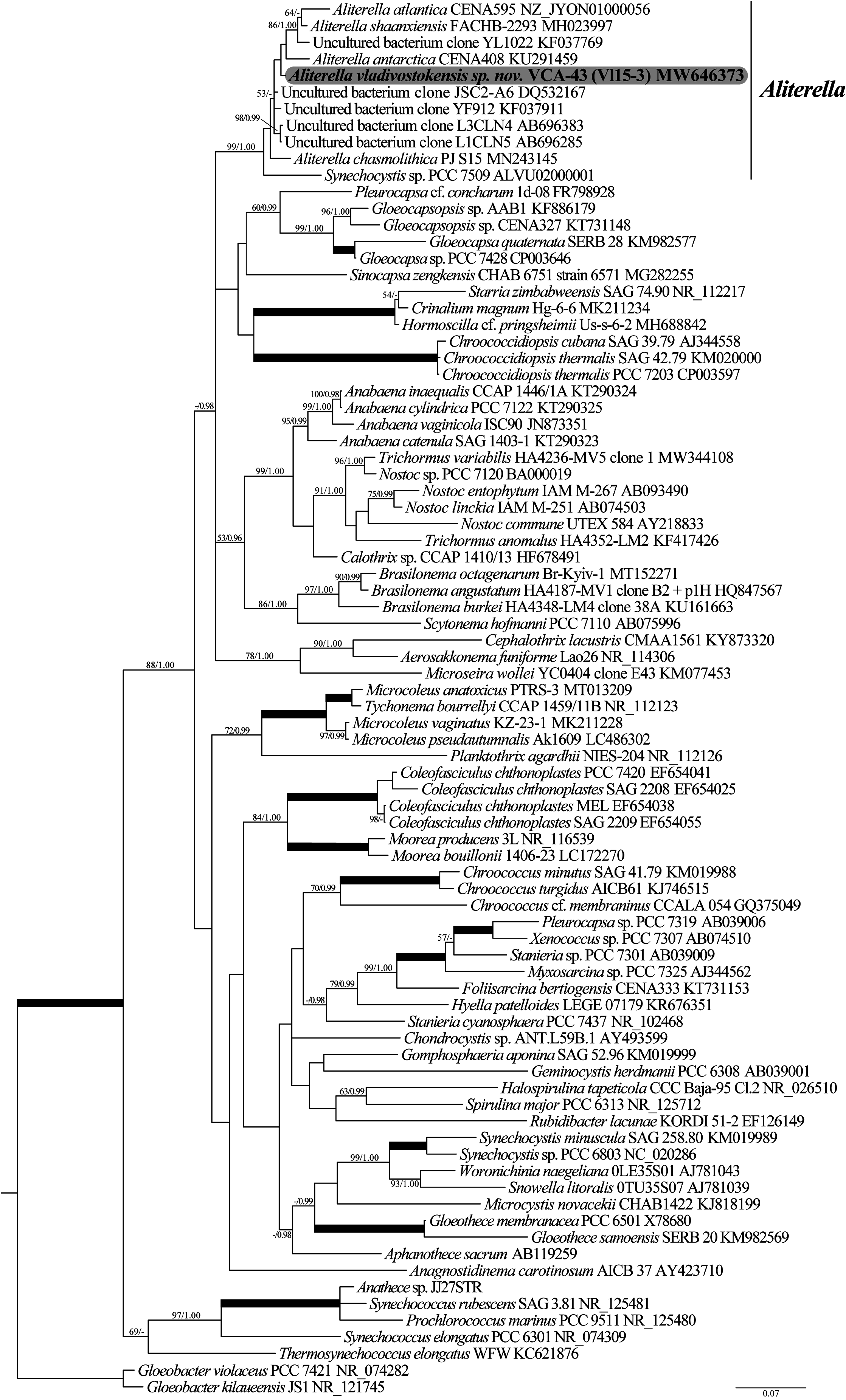
Phylogenetic analyses (ML and BI) clearly indicated that A. vladivostokensis was a member of the strongly supported (99/1.00; BP/PP; Fig. 2 View FIGURE 2 ) Aliterella generic clade. Branching pattern between the clade members remained largely unresolved ( Fig. 2 View FIGURE 2 ).
We observed the highest 16S rRNA sequence similarities based on p -distance analysis between our isolate and an uncultured bacterial clone (DQ532167) isolated from clean spacecraft assembly rooms where spacecrafts are assembled (99.5%; Table 1 View TABLE 1 ). A. antarctica ( KU291459 View Materials ) and A. shaanxiensis ( MH023997 View Materials ) were the most similar to each other among described Aliterella species , (99.1%). Since no ITS sequences were available for the uncultured bacterial clones resolved as members of the Aliterella clade (5 accessions), we only compared the percentage of sequence dissimilarity between aligned 16S–23S ITS regions for the described taxa and Synechocystis sp. PCC 7509 (6 accessions). The dissimilarity varied from 6.4% to 17.5% (11.6%–17.5% between A. vladivostokensis and the other species; Table 2 View TABLE 2 ).
Comparison of ITS secondary structures of the D1–D1′ and Box-B helices showed that they have similar patterns of bulges and terminal loops ( Fig. 3 View FIGURE 3 , 4 View FIGURE 4 ). The D1–D1′ helix was almost invariant in length (65–67 bp) and structure with 42 conserved sites among Aliterella species ( Fig. 3 View FIGURE 3 ). In Synechocystis sp. PCC 7509 the helix was shorter (57 bp) due to the deletion in the terminal loop ( Fig. 3 View FIGURE 3 ). The basal part of the D1–D1′ helix consisted of a conservative 4-bp double-stranded region followed by the internal (bilateral) loop (positions 5–6 and from 55–57 to 61–63 in different Aliterella species ; 5–6 and 47–53 in Synechocystis sp. PCC 7509) and side loop with a single unpaired base (position 51–52, position 43 for Synechocystis sp. PCC 7509). Substitutions in the bilateral loop (base change U → C at position 57) and in the side loop (A → C at position 51) differentiated A. vladivostokensis . The terminal loop of the helix consisted of five bp and also harbored a unique marker mutation for the new species (A → C at position 29). Most species had two internal loops in the central part of D1–D1′ helix with the exception of A. shaanxiensis , which had three loops and Synechocystis sp. PCC 7509 with the one loop. We found one compensatory base change (CBC) and five hemi-compensatory base changes (hCBCs) in our secondary structure models. While CBC (A-U → G-C) was shared between A. vladivostokensis and A. antarctica (at positions 19–42 in both cases), the hCBCs were attributed to A. atlantica (C-G → U-G, at position 22), A. antarctica (two substitutions C-G → U-G at positions 20 and 27), A. chasmolithica (G-C → G-U, at position 33; Fig. 3 View FIGURE 3 ), and Synechocystis sp. PCC 7509 (C-G → U-G at position 13).
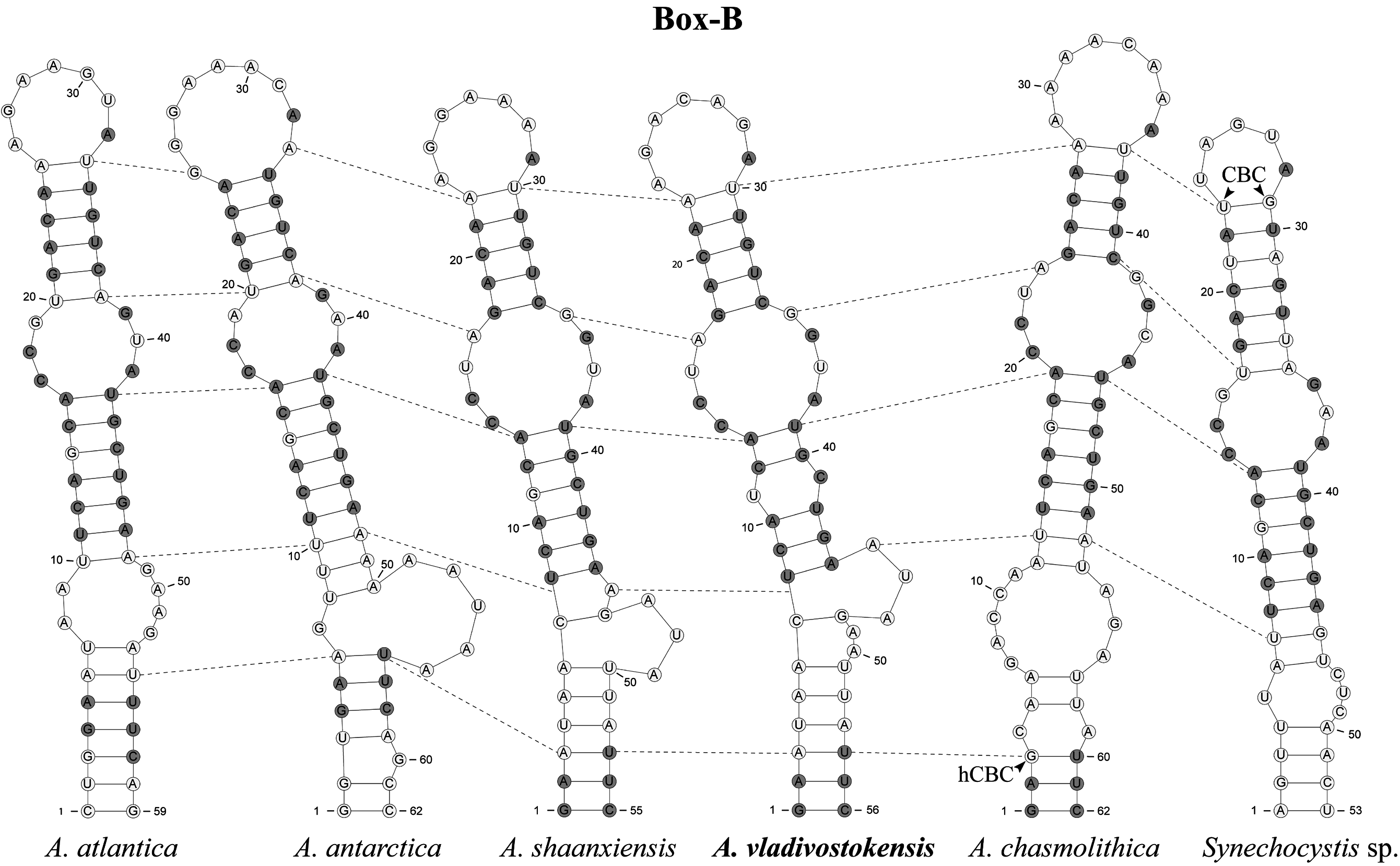
The Box-B helix was more variable in both primary and secondary structure (53–62 bp, 29 conservative sites); therefore, homologous nucleotides were difficult to identify in some cases ( Fig. 4 View FIGURE 4 ). The middle part of the Box-B helix had two ( A. atlantica ) to four ( A. vladivostokensis ) mostly bilateral internal loops. Point mutations, mostly found in single stranded domains, and indels frequent in the basal part of the Box-B altered the bulges location between species. A base substitution G → U at position 11 in the Box-B disrupted nucleotides pairing that led to an additional internal loop formation in A. vladivostokensis sequence ( Fig. 4 View FIGURE 4 ). The Box-B helix in Synechocystis sp. PCC 7509 differed from those in other species in lacking conservative motive at the base of the helix and shorter terminal loop (5 vs. 7–9 bases; Fig. 4 View FIGURE 4 ). We found one CBC in Synechocystis sp. PCC 7509 (A-U → U-G, at positions 3 and 29) and one hCBC in A. chasmolithica (A-U → G-U, at position 3).


| R |
Departamento de Geologia, Universidad de Chile |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |