
Palaeictops altimontis, Velazco & Novacek, 2016
|
publication ID |
https://doi.org/ 10.1206/3867.1 |
|
DOI |
https://doi.org/10.5281/zenodo.5056601 |
|
persistent identifier |
https://treatment.plazi.org/id/0386879B-FB16-9C65-FE41-31E888E9FDBA |
|
treatment provided by |
Felipe |
|
scientific name |
Palaeictops altimontis |
| status |
sp. nov. |
Palaeictops altimontis , new species
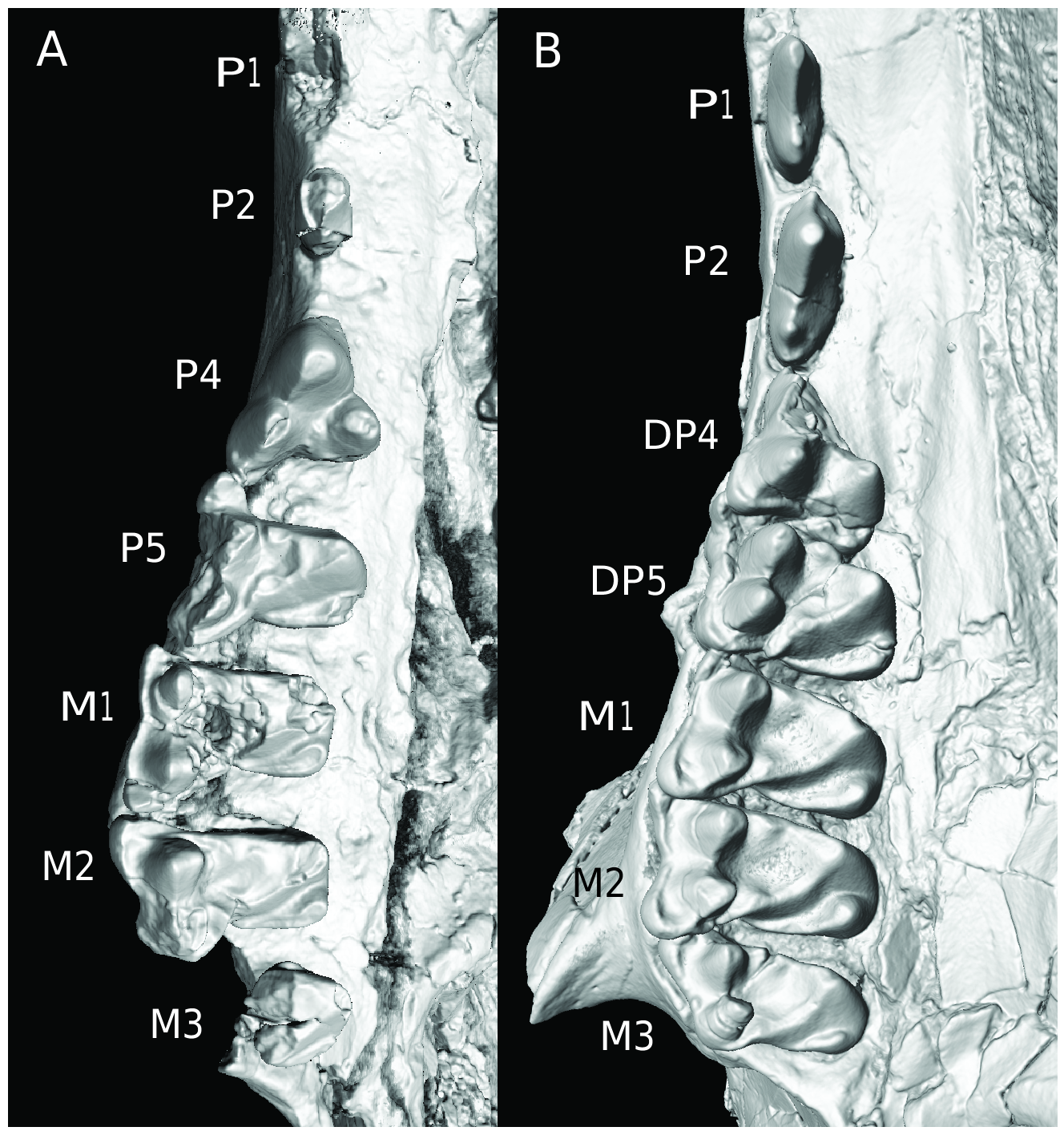
Figures 5B View FIG , 13–15 View FIG View FIG View FIG , table 1
Palaeictops sp. Novacek, 1977: 21.
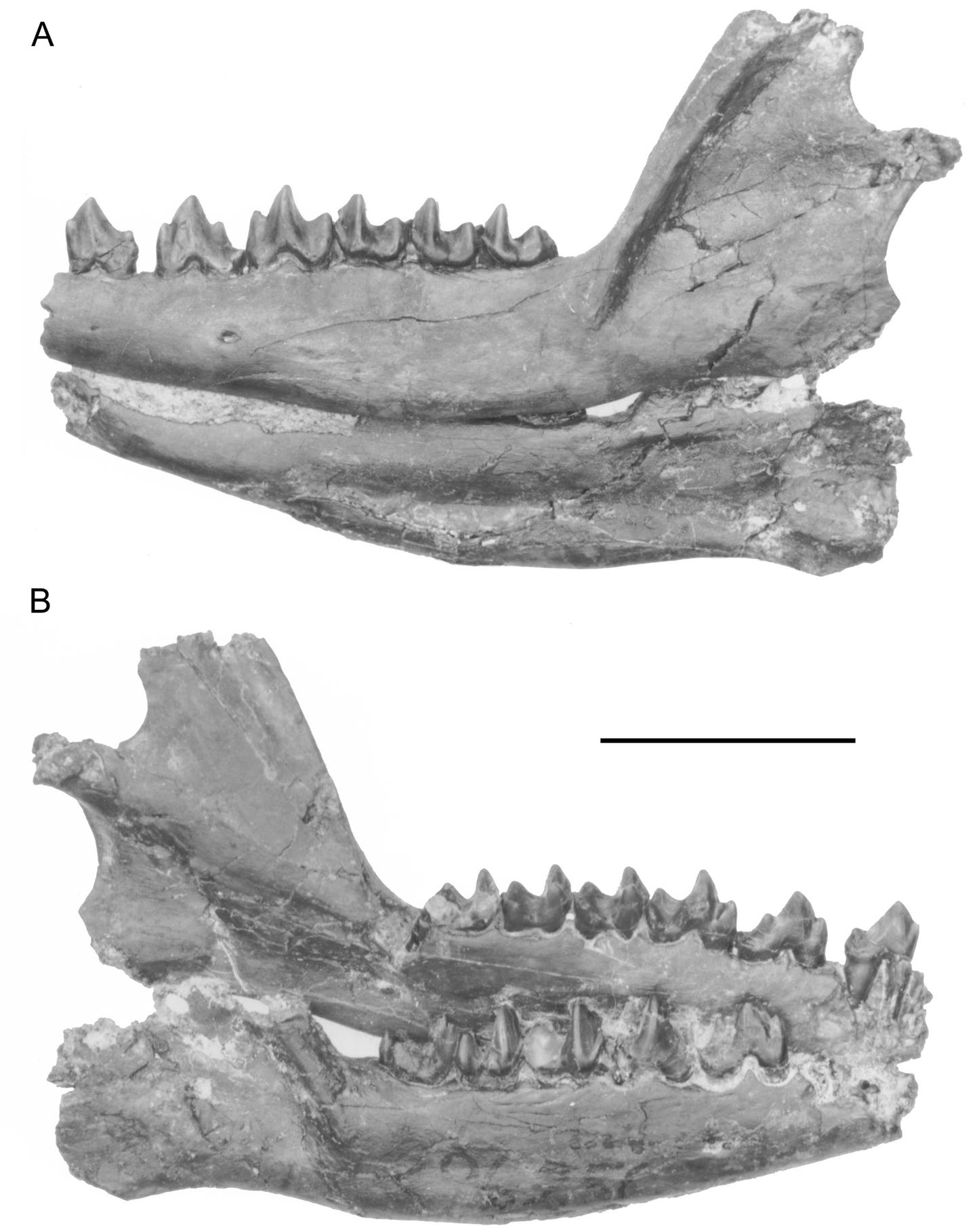
HOLOTYPE: AMNH 96250 About AMNH (figs. 5B, 13B), consisting of a nearly complete skull that retains P1–P2, DP4–DP5, M1–M3 on the left side; the right side retains P5–M3. Left ramus with p2, and p4–m3 (fig. 15A–B). Right ramus with p4, p5 (damaged), and m1–3 (fig. 15B).
REFERRED SPECIMENS: AMNH 88400, right ramus with p4–5; AMNH 101955, left ramus with P2, P5; AMNH 105032, left ramus with p4, m3; AMNH 99301, left maxilla with P4–M3; AMNH 113880 About AMNH , left ramus with p4–m3. All from the type locality .
TYPE HORIZON AND LOCALITY: Unit 24 (bone bed A) East Fork Basin, northeast of Dubois, Freemont Co., Wyoming, about 500 feet above the local base of the Tepee Trail Formation (see McKenna, 1980: 337), Middle Eocene (Uintan NALMA).
ETYMOLOGY: From the Latin: altus, “high,” and mons, “mountain.” Refers to the dramatic montane settings of the type locality (see Love, 1939; McKenna, 1980).
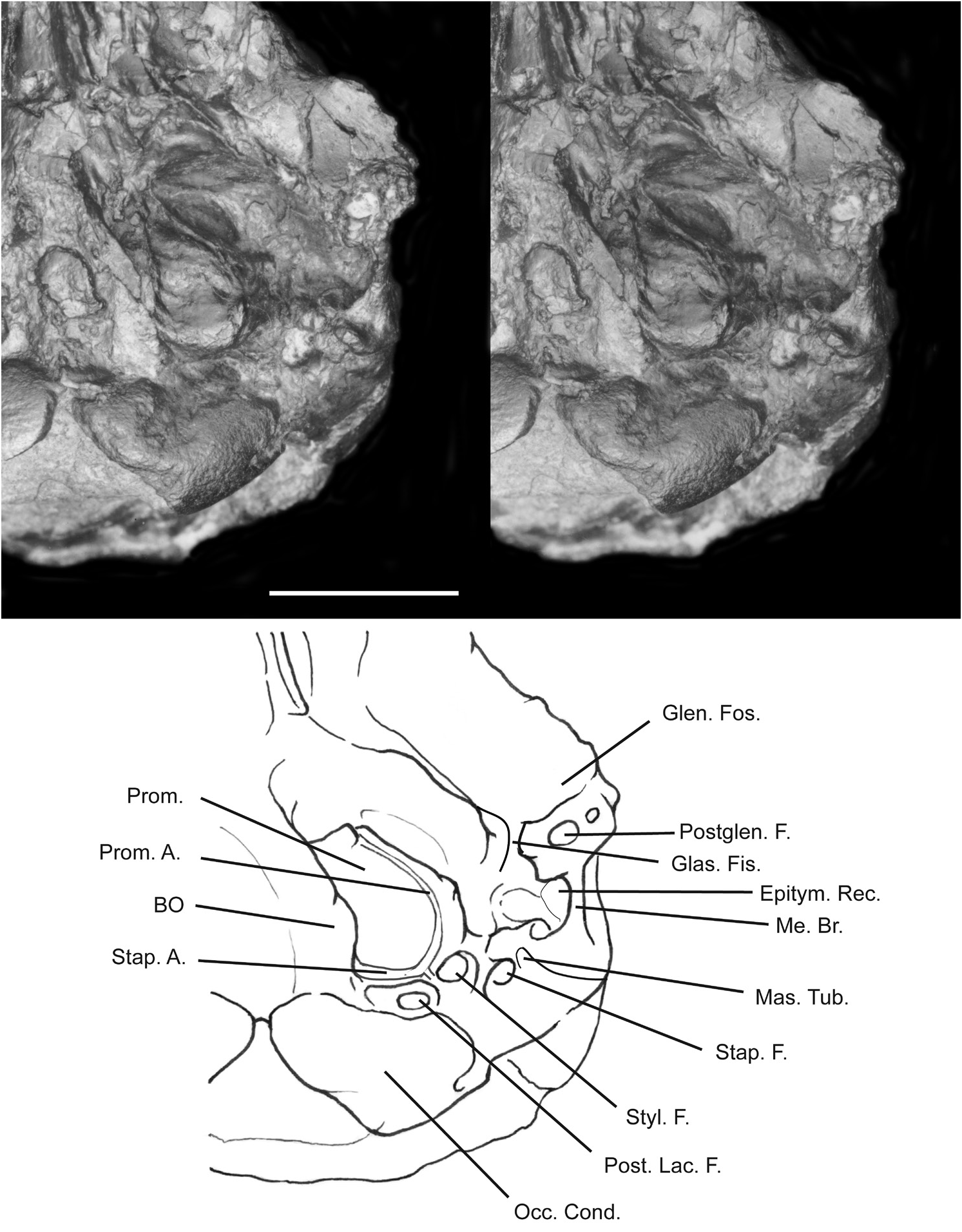
DIAGNOSIS: Like Palaeictops bridgeri in having p5 with open prefossid and shortened talonid, but differs in smaller size, in lacking an anterior accessory cuspule at labial base of paraconid; p5 metaconid aligned less obliquely relatively to protoconid; more anteroposteriorly compressed trigonid on m2; and less transverse M2 with a deeper ectoflexus. Differs from P. bicuspis , P.multicuspis , and P. matthewi in having smaller, more anteriorly separated paraconid on p5 and lower trigonids on m1–3. Skull like P. bicuspis but unlike Leptictis in having single sagittal crest (fig. 13A). Skull like Leptictis but unlike P. bicuspis in having posteriorly narrow nasal elements. Basicranium differs from Leptictis (cf. fig. 14 and Novacek, 1986: fig. 22) in having: (1) a postglenoid foramen positioned more laterally and farther from the anteroexternal edge of the tympanic cavity; (2) a more transversely flared basioccipital that overlaps ventrally the promontorium of the petrosal; (3) shallower grooves on the promontorium for the promontory and stapedial branches of the internal carotid artery; (4) a posterior lacerate foramen only slightly larger than the stapedius fossa (much larger and more oval than stapedius fossa in Leptictis ); (5) a cochlear fossula with a more expanded, dorsal rim; (6) a narrower bridge of the mastoid tubercle of the petrosal extending from ventral rim of the cochlear fossula between the stylomastoid foramen and stapedius fossa; (7) a paraoccipital process that is less extensive, so that the distance is shorter between stylomastoid foramen and posterior margin of basicranium; and (8) a shallower groove on paraoccipital process for the digastric muscle.
DESCRIPTION AND COMPARISONS: The front of the skull of Palaeictops altimontis (AMNH 96250) is badly damaged (fig. 13A–B), and the premaxillae are not preserved, except for a small nasal process on the right side of the skull. Most of the paired nasal elements can be seen in dorsal view (fig. 13A). Posteriorly the nasals are narrow, as in Leptictis . However, unlike the latter, the contact of the nasal and the frontal is along a more obliquely oriented suture (fig. 13A). It is difficult to ascribe much taxonomic significance to this difference, as the form of this suture varies to some degree in Leptictis . On the right side of the skull, the maxilla has been displaced laterally, and the contact of this element with the nasal is marked by a faint ridge that divides the dorsal (horizontal) process of the nasal from a more vertically oriented process (fig. 13A). From this, it is clear that the maxilla overlaps the nasal for a considerable extent on the skull roof and nasalfacial exposure (fig. 13A). Differential growth of the maxilla and nasal may thus account for the relatively narrowed nasal exposure in Palaeictops and Leptictis .
The maxilla has a broad contact with the frontal along an oblique suture (fig. 13A). The condition resembles that in Leptictis . In Palaeictops bicuspis this contact is narrower, because the posterior nasals are much broader in their contact with the frontals (fig. 2A). The antorbital fossa is distinct, though shallower than in Leptictis . The ventral border of the fossa is marked by a pronounced ridge. The anterior foramen of the infraorbital canal (preserved only on the right side of the skull) is a circular, but somewhat smaller, opening than in the Leptictis (fig. 13A–B). As in the latter, the infraorbital canal is relatively short in length; it opens above M1 (fig. 13A–B). Unfortunately, the important relationships of the maxilla with other elements of the orbital region cannot be seen due to poor preservation.
The palatine has a basically similar construction as that in Leptictis . Its posterior margin (coincident with the posterior margin of the palate) lies between the last molars (fig. 13B). The margin is biconcave with a distinct, rounded postpalatine torus (fig. 13A–B). The minor palatine foramen is a large opening in the pars perpendicularis that meets the pterygoid (fig. 13A– B). The dorsally trending route of the minor palatine foramen cannot be traced as orbital foramina are obscured by damage. It appears, however, that the dorsal exit of the minor palatine foramen and the sphenopalatine foramen were in closed proximity within a shallow depression, a condition also seen in Leptictis .
The lacrimal is a small triangular element vaguely demarcated on both sides of the skull (fig. 13A–B). As in Leptictis , the lacrimal foramen is confined fully within the orbit. It faces posteriorly and lies directly below the dorsal ridge of the lacrimal (fig. 13A–B). A lacrimal tubercle is present, though it appears much weaker than this structure in Leptictis .
The jugal is not preserved, but a broken, rugose surface on the anterior root of the zygoma suggests that this element contacted the lacrimal (a common and probably primitive eutherian trait seen in other leptictids).
The frontal is poorly demarcated in the orbit, so it is impossible to determine whether it was isolated from the orbital process of the maxilla by the intrusive palatine. Its separation from the maxilla, however, seems likely, because the palatine-maxillary boundary lies directly above the pars perpendicularis of the palatine. This relationship is like that in Leptictis . Accordingly, it’s probable that Palaeictops shared with the latter the same orbital mosaic. A small ethmoidal foramen is preserved near the frontal-orbitosphenoid suture in the right side of the skull (fig. 13A–B).
The most notable feature of the skull roof is the single sagittal crest, which is best exposed in the posterior parietal near the junction of the sagittal and lambdoidal crests (fig. 13A). More anteriorly, bone is missing and the brain endocast underneath shows the medial longitudinal fissure (fig. 13A). In addition to having only a single median sagittal crest, this skull differs from Leptictis in showing much less sculpturing for attachment of the temporalis muscle on the parietal and squamosal. This, in combination with the comparatively smaller postglenoid process, suggests a weaker development of the temporalis complex of the jaw-closing apparatus. As noted in Novacek (1986), Leptictis shows a strong emphasis of orthal shear.
Another interesting feature of the skull roof in Leptictis is the extension of the parietal around the lambdoidal crest and its exposure as a small triangular process on the occiput. On the left side of the skull of Palaeictops altimontis , there is a break that may indicate the boundary between the parietal and interparietal, suggesting that the parietal in this form also developed an occipital process. Unfortunately, because of poor preservation, this rather anomalous mammalian condition is not clearly identified in Palaeictops .
Features of the orbitosphenoid and alisphenoid in the orbit are not clearly preserved (fig. 13B). The presphenoid appears to have a ventral median keel as in Leptictis . On the left side of the skull, there appears to be a short, but badly damaged alisphenoid canal just anterior to the foramen ovale.
The pterygoid shows prominent, vertical, entopterygoid crests that form the medial walls of the ectopterygoid fossae (fig. 13A–B). Although this region is damaged, there is evidence for the presence of lateral ectopterygoid crests of the alisphenoid, indicated by the extensive development of the fossae and the flaring of a ridge on the right side of the skull that lies lateral to the entopterygoid crests (fig. 13A–B). As noted in Novacek (1986: 45) it is probable that the manner in which the internal pterygoid muscles originated from the skull is quite similar in Palaeictops and Leptictis .
The basisphenoid is closely fused with adjacent elements. In ventral view it forms a trapezoidal platform with weak sculpturing for pharyngeal grooves and rectus capitis muscles (fig. 13A–B).
The squamosal is strongly distorted by the dorsoventral flattening of the skull (fig. 13A). However, a few features of interest are recognizable. As noted above, there is no marked rugosity or ornamentation of the lateral moiety of the squamosal in the temporal region. Unlike Leptictis the suprameatal fossa behind the zygomatic process of the squamosal is quite shallow. A small opening lies directly above this fossa on the left side of the skull. This is likely the suprameatal foramen. In Leptictis , the suprameatal foramen is much larger and is situated more ventrally, well within the deep suprameatal fossa. The glenoid fossa for articulation with the lower jaw is a shallow, broad surface, somewhat more extensive and less concave than in Leptictis (fig. 14). Although the skull on both sides is damaged in the region of the postglenoid process, it is clear that this feature is smaller than in Leptictis . There is only a very narrow bridge of bone representing the root of the postglenoid process. Furthermore, the postglenoid foramen lies posterior and slightly lateral to this area (fig. 14). In Leptictis the foramen has shifted to a more medial position, presumably repositioned by the marked expansion of the swollen postglenoid process.
The tympanic region of AMNH 96250 is remarkably well preserved and, thus, shows much more detail than other regions of the skull. There are some obvious departures from the condition in Leptictis . The promontorium of the petrosal is more smoothly rounded and does not taper as distinctively in its anterior region (cf. fig. 14 and Novacek, 1986: fig. 22). The regularity in surface features of the promontorium is the result of two other characteristics. The sulci for the promontory (= internal carotid) and stapedial arteries are very narrow and faint, whereas in Leptictis they form distinctly deep troughs (cf. fig. 14 and Novacek, 1986: fig. 22). In addition, the petrosal crest that arcs on the medial surface of the promontory is only a weak flange, whereas in Leptictis it develops as a distinct and prominent ridge with a rugose contact surface with the entotympanic bulla. Despite the weaker petrosal crest in Palaeictops altimontis , it is likely that this form also had an entotympanic bulla. Although this structure is not present in the specimen, it is also commonly not preserved in skulls of Leptictis , so it seems to be a feature readily lost during burial. Another feature of the promontorium peculiar to P. altimontis is the very broad development of the dorsal lip of the cochlear fossula (fig. 14), which overhangs the fenestra rotunda. In Leptictis this feature is also distinct but is less prominent.
There is, as in Leptictis , a distinct glaserian fissure in the lateral tympanic roof of AMNH 96250. Lateral to the region of the excavated facial canal, there appears to be an epitympanic recess, although its size is uncertain, because the meatal bridge of the squamosal has collapsed (fig. 14). This bridge, the suprameatal surface of the squamosal, is much narrower than in Leptictis (cf. fig. 14 and Novacek, 1986: figs. 20, 21). Moreover, the postympanic process, which forms the posterior buttress for the roof of the meatus, is weaker in Palaeictops . Behind this buttress, the paraoccipital process of the petrosal is somewhat broader in exposure, and it lacks the distinctive groove for the digastric muscle seen in Leptictis .
Posterior to the facial canal is a very well-defined stylomastoid foramen. This opening seems even somewhat larger than in Leptictis , though the difference here is of dubious significance. In the left tympanic region, there is a small process that arises from the mastoid just medial to the postympanic process and extends below the facial canal just short of contact with the promontorium in the region of the fenestra vestibuli. This process is likely that of the mastoid tubercle, which may also represent the fusion of the tympanohyal with the petromastoid (fig. 14). The cup-shaped ventral depression on this tubercle characteristic of Leptictis is not present, although the tubercle is so badly damaged that presence of this fossa in Palaeictops cannot be ruled out. The posterior lacerate foramen is, as in Leptictis , merged with the jugular foramen, so there is only one exit for the internal jugular vein and cranial nerves IX, X, and XI. Because the ventral lip of the cochlear fossula is so expanded, there is no narrow trough between the stapedius fossa and the posterior lacerate foramen (fig. 14). This latter feature is distinctly present in Leptictis .
The occiput in Palaeictops altimontis is badly compressed and damaged, but it is apparent that there was a prominent mastoid exposure in this region. The anterior edge of the ventral occipital condyles has the sigmoid curvature seen in Leptictis and lipotyphlans.
The mandible is deepest below m1 (fig. 15). There is a small mental foramen located below p2 and below the posterior root of p4. At the back of the jaw the masseteric fossa is well excavated. The coronoid process shows some tapering dorsally (fig. 15), but does not show the extreme posterior concavity seen in Leptictis (figs. 15 and Novacek, 1986: fig. 1). Because the extremity of the coronoid process is missing, it is uncertain whether the process had a hooklike outline as in Prodiacodon tauricinerei or a more bluntly round curved process as in Palaeictops robustus (fig. 17). The angular process is damaged in both right and left mandibles. The articular surface of the jaw condyle is slightly broader medially than laterally. The surface features of the condyle are poorly preserved.
There are no upper incisors or canines preserved in AMNH 96250 (fig. 13). The canine alveolus is present on the right side of the skull. P1 is bicuspid, single rooted, and triangularly shaped in lateral outline. P2 is tricuspid, double rooted, and triangularly shaped in lateral outline. The central cusp of P2 is dominant while the most anterior cusp is very minute (fig. 5B).
AMNH 96250 shows a remarkable condition, wherein the right DP4–5 are present and only moderately worn, and the left P5 is already erupted (left P4 is missing) (figs. 5B, 13B). DP4 is roughly triangular in occlusal view with well-developed, somewhat inflated paracone, metacone, and protocone (fig. 5B). There is also a crenulated postcingulum. DP5 is molariform with well-developed paraconule, metaconule, and postcingulum in addition to three inflated main cusps (fig. 5B). There is a pronounced parastylar spur that is overlapped ventrally by the metastylar spur of DP4 (fig. 5B).
A P4 is preserved in AMNH 99301, but it is badly worn. The tooth is roughly triangular in occlusal view, although its parastylar spur is very strong, as is typical of leptictids. P5 is fully molariform with distinct conules. The hypocone is also well developed to nearly a third the height of the protocone. P5 has a strong parastylar spur, but the labial margin of the tooth shows no appreciable invagination.
The upper molars are essentially like P5 although they have broader crowns, more inflated cusps and conules, and (except for M3) relatively larger hypocones and talon basins (fig. 5B). The general construction of the dentition is very like that in other species of Palaeictops and in Leptictis . The labial margins of the upper molars have shallow inflexions in contrast to the condition in Prodiacodon . There is no evidence of a doubling of the paraconule as in the upper molars of Prodiacodon (fig. 5B; cf. Novacek, 1986: fig. 4B).
Lower incisors, canine, and p1 are missing from all specimens (fig. 15). The p2 is elongate, trenchant, and two rooted, with a small anterior cuspule, a large central cusp, and a low heel (fig. 15). The p4 resembles p2 but has four cusps: a small anterior cuspule, a large “central” cusp, a small cuspule on the posterior ridge of the central cusp, and a low cuspid heel (fig. 15). The p5 is molariform with a moderately developed paraconid well separated from the metaconid and protoconid and has an elongated talonid basin bordered by three cusps. The crista oblique contacts the posterior wall of the trigonid nearly directly below the posterior trigonid notch. As noted above the p5 resembles Palaeictops bicuspis in having a very open trigonid due to the salient paraconid. The metaconid is less oblique in its alignment with the protoconid than in P. bicuspis . The lower molars are very characteristic of Palaeictops and Leptictis as a whole (see Novacek, 1986: 17), although the trigonid of m2 is notably compressed due to the crestiform paraconid.
REMARKS: The upper cheek teeth of Palaeictops altimontis resemble more closely the corresponding teeth of Leptictis than any other Paleocene or Eocene leptictid. Palaeictops altimontis lacks, however, the distinctive trenchant p4 of Leptictis , a condition more closely approached in P. matthewi . In addition, P. altimontis retains the single sagittal crest and several basicranial characters that are clearly modified in Leptictis . Aspects of molar morphology are specializations that separate all known species of Palaeictops from Leptictis .
Palaeictops bridgeri and P. altimontis are thus far known only from Middle Eocene assemblages. They depart from Early Eocene Palaeictops species in the structure of the p5 paraconid.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Leptictinae |
|
Genus |
Palaeictops altimontis
| Velazco, Paúl M. & Novacek, Michael J. 2016 |
Palaeictops
| Novacek, M. J. 1977: 21 |