
Diploneis cristata, Jovanovska & Wilson & Hamilton & Stone, 2023
|
publication ID |
https://doi.org/10.11646/phytotaxa.593.1.1 |
|
DOI |
https://doi.org/10.5281/zenodo.7875103 |
|
persistent identifier |
https://treatment.plazi.org/id/038487E2-FFD1-264D-BCF1-FF09B9B67630 |
|
treatment provided by |
Plazi |
|
scientific name |
Diploneis cristata |
| status |
sp. nov. |
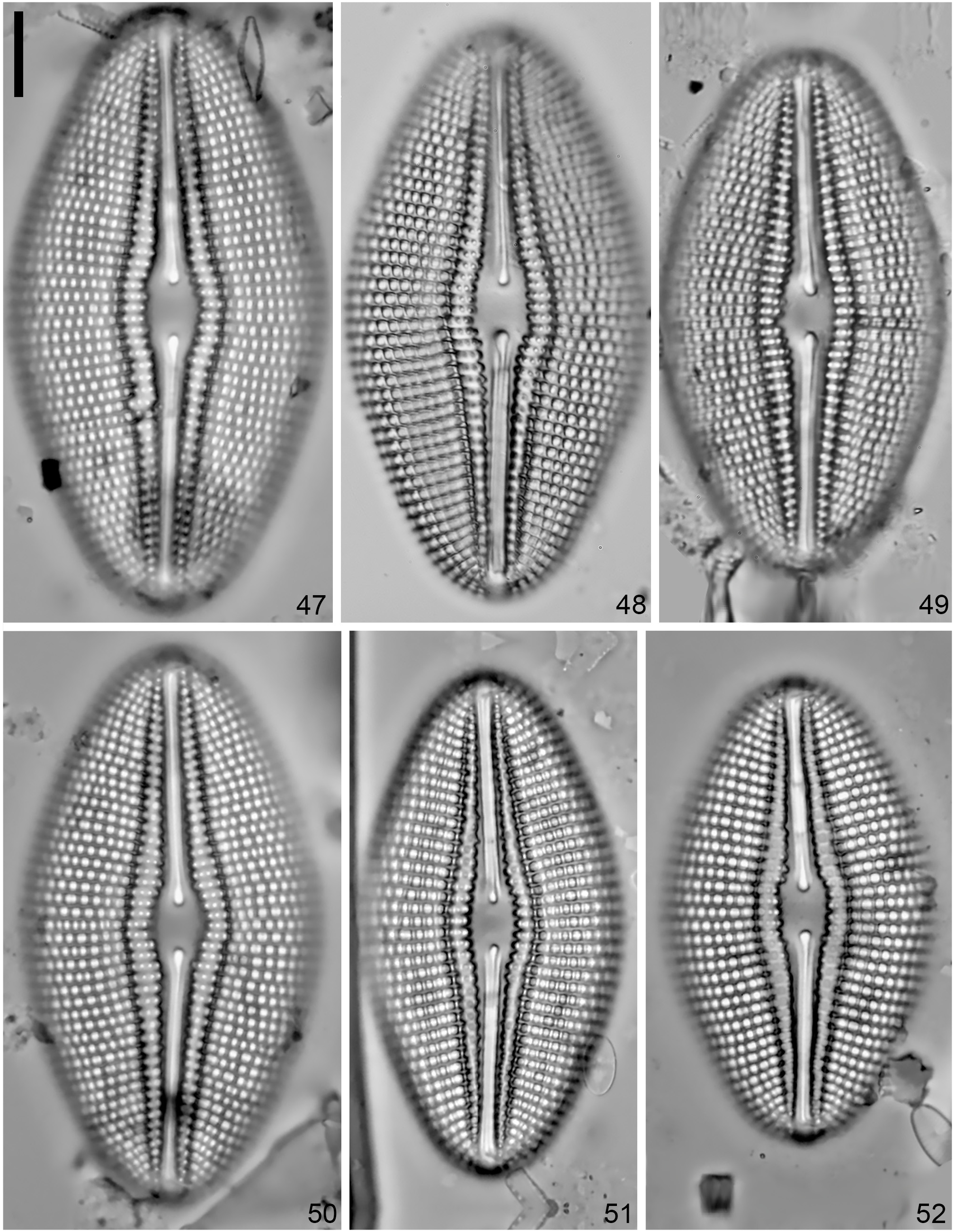
Diploneis cristata sp. nov. (LM Figs 47–52 View FIGURES 47–52 , SEM Figs 53–60 View FIGURES 53–56 View FIGURES 57–60 )
Valves are weakly asymmetric, rhombic-elliptic to elliptic with convex margins and bluntly round apices ( Figs 47–53 View FIGURES 47–52 View FIGURES 53–56 ). Valve length is 47–80.5 μm and valve width is 26–41 μm. The axial area is narrow, lanceolate, widening at the center to form a lanceolate central area ( Figs 47 View FIGURES 47–52 , 55 View FIGURES 53–56 ), 4.5–8.5 μm wide. Externally, the longitudinal canal is broad, lanceolate to linear, slightly expanded in the middle of the valve with three to four rows of cribrate (10–20 poroids) areolae narrowing into one at the valve apices ( Figs 53, 55, 56 View FIGURES 53–56 ). Internally, a thick non-porous slightly raised silica plate encloses the longitudinal canal ( Figs 57, 59 View FIGURES 57–60 ). Externally, the raphe is filiform, curved; the proximal ends are weakly deflected and positioned within a teardrop depression ( Figs 53, 55 View FIGURES 53–56 ). The distal raphe ends are unilaterally bent to the same side and terminate on the valve face ( Figs 53, 56 View FIGURES 53–56 ). Internally, the raphe branches are slightly arched with simple proximal and distal ends that are slightly elevated in a deep depression formed by the longitudinal canal ( Fig. 57 View FIGURES 57–60 ). The striae are parallel at mid-valve becoming radiate towards the valve apices, 7–8 in 10 μm. Striae are uniseriate throughout ( Fig. 54 View FIGURES 53–56 ). The striae are composed of large round to rectangular areolae covered externally with cribra (>40 poroids; hardly visible on the illustrated specimens), 7–8 in 10 μm. Each areola opens into deep pits and sometimes separated by short crested fin-like ridge ornamentations that are forming from the areolae wall (white arrowed Fig. 54 View FIGURES 53–56 ). The inter-areolar thickenings have fin-like shaped silica ornamentations serrated into ca. 3–9 notched edges ( Figs 54, 56 View FIGURES 53–56 ). The areolae increase in size towards the valve margins ( Figs 53, 55 View FIGURES 53–56 ). Internally, the alveoli open via a single elongated opening covered with a thin silica layer ( Figs 59, 60 View FIGURES 57–60 ). The valvocopula has serrated advalvar edges ( Figs 57, 59, 60 View FIGURES 57–60 ).
Type:— REPUBLIC OF ZAMBIA, Lake Tanganyika , Chituta Bay, at 768 m elevation; mud and mussels, 33 m water depth, collected SCUBA diving, 8°42’56.0” S 31°09’15.0” E, W. Salzburger, 5 th September 2018 ( holotype designated here, circled specimen BM-108995! = Fig. 48 View FIGURES 47–52 , GoogleMaps isotypes ANSP-GC17224 !, CANA-129321!). Type material CANA-129321. Registration: http://phycobank.org/103701 GoogleMaps
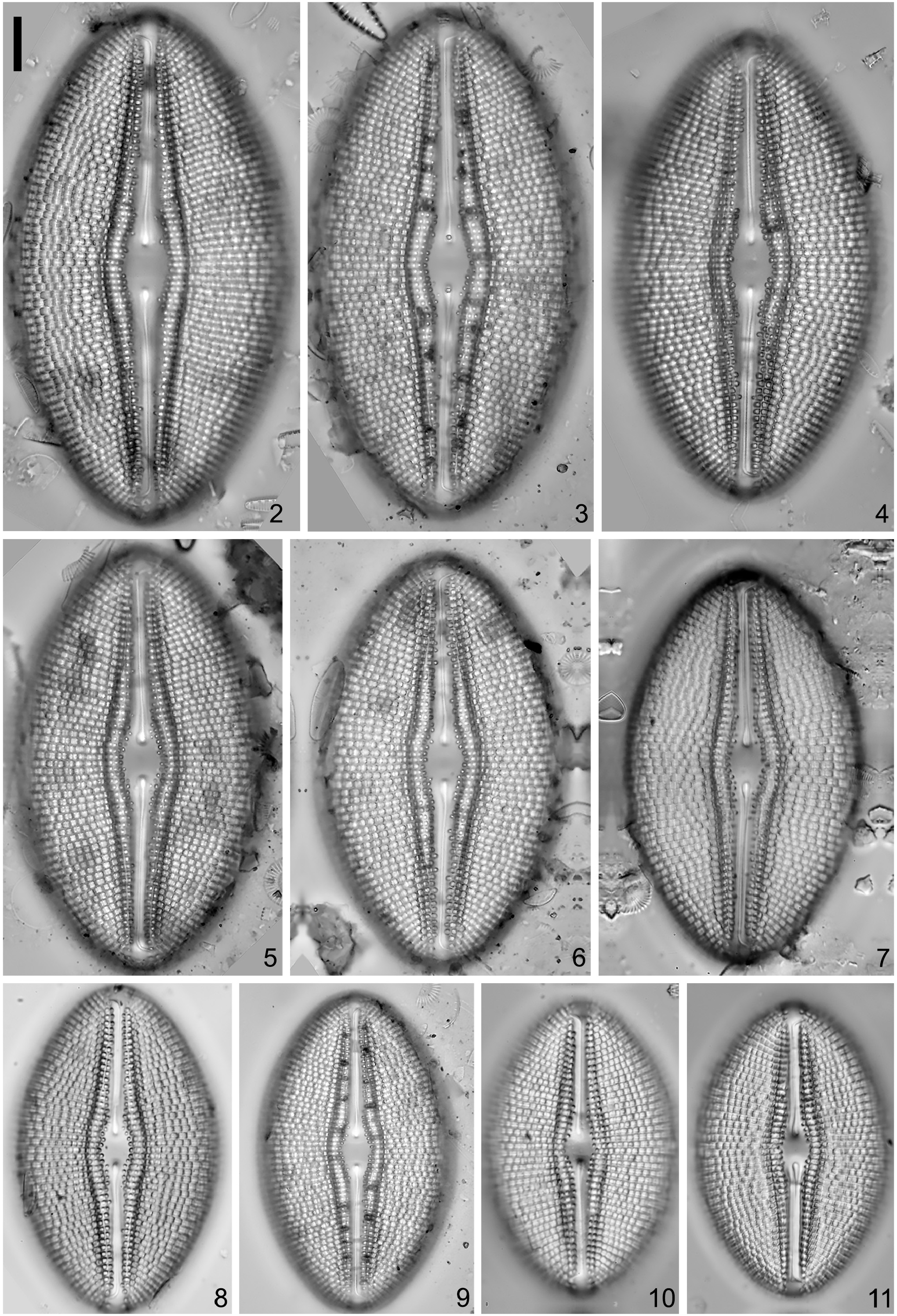
Pictures of the isolated specimen:— LM micrograph on 1000× magnification ( Fig. S2e View FIGURES 2–11 ).
Sequence data:— Plastid gene rbc L sequence (GenBank accession: OQ 660279).
Etymology:— The specific epithet ‘ cristata ’ is derived from the distinct crested fin-like ridge ornamentations on the valve face.
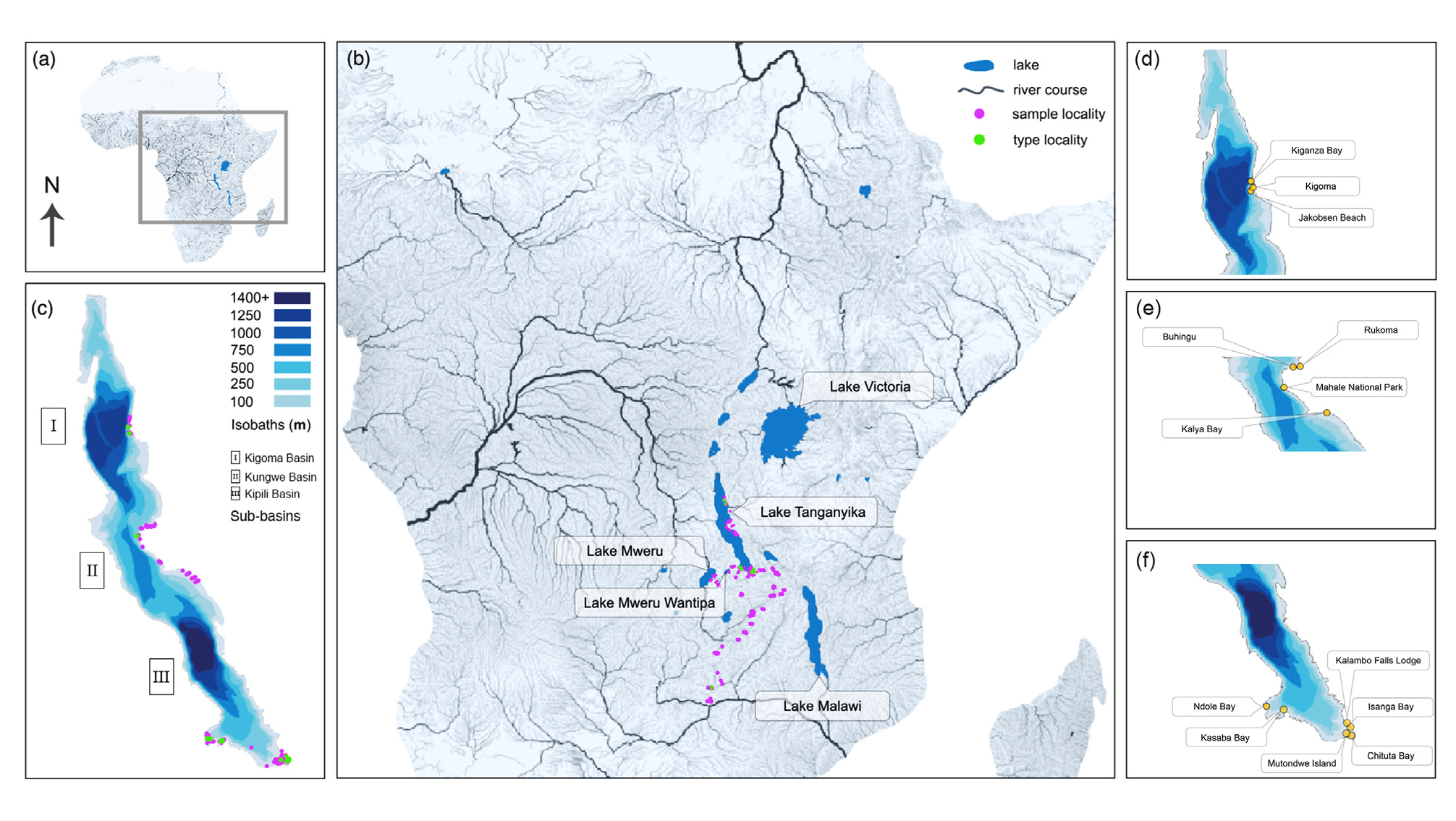
Ecology and distribution:— Diploneis cristata sp. nov. has been observed along the Tanzanian and Zambian coasts of Lake Tanganyika. It is not a very frequent species in this alkaline lake with moderate mineral content, occurring mainly on muddy and sandy substrates between 5 and 25 m water depth, but also as tychoplankton in the littoral areas and in plankton up to 60 m depth. This species occurs mainly in the southern sub-basin at Kalambo Falls Lodge, Isanga Bay, Chituta Bay, Mutondwe Island, and Ndole Bay ( Fig. 1c, f View FIGURE 1 ). In this sub-basin, the species occurs mainly together with D. gigantea sp. nov., D. fossa sp. nov., D. cocquytiana sp. nov., and D. salzburgeri sp. nov. Smaller population sizes of this species have been observed in the central and northern sub-basins in Kalya Bay, Mahale National Park, Rukoma area, Buhingu Island, and Kiganza Bay ( Fig. 1c–e View FIGURE 1 ). In these areas, the species usually coexists with D. serrulata sp. nov., D. cocquytiana sp. nov., D. lunata sp. nov., D. salzburgeri sp. nov., and D. tanganyikae sp. nov.
Main differential characters:— Valve size and shape, striae density, external thick fin-like ornamentations across the valve, fin-like thickening within areolae, and poroids>40 per areola.
Similar species:— Diploneis tanganyikae sp. nov., D. salzburgeri sp. nov., and Diploneis buriatica Kulikovskiy & Lange-Bertalot (2015: 91) .


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |