
Dinothenarus Thomson, 1858
|
publication ID |
https://doi.org/10.1206/0003-0082(2000)287<0001:ROTNTT>2.0.CO;2 |
|
persistent identifier |
https://treatment.plazi.org/id/0384878F-D830-FB3E-FF21-694E0B04FB04 |
|
treatment provided by |
Carolina |
|
scientific name |
Dinothenarus Thomson, 1858 |
| status |
|
Dinothenarus Thomson, 1858 View in CoL ( stat. nov.)
TYPE SPECIES: Staphylinus pubescens DeGeer, 1774 .
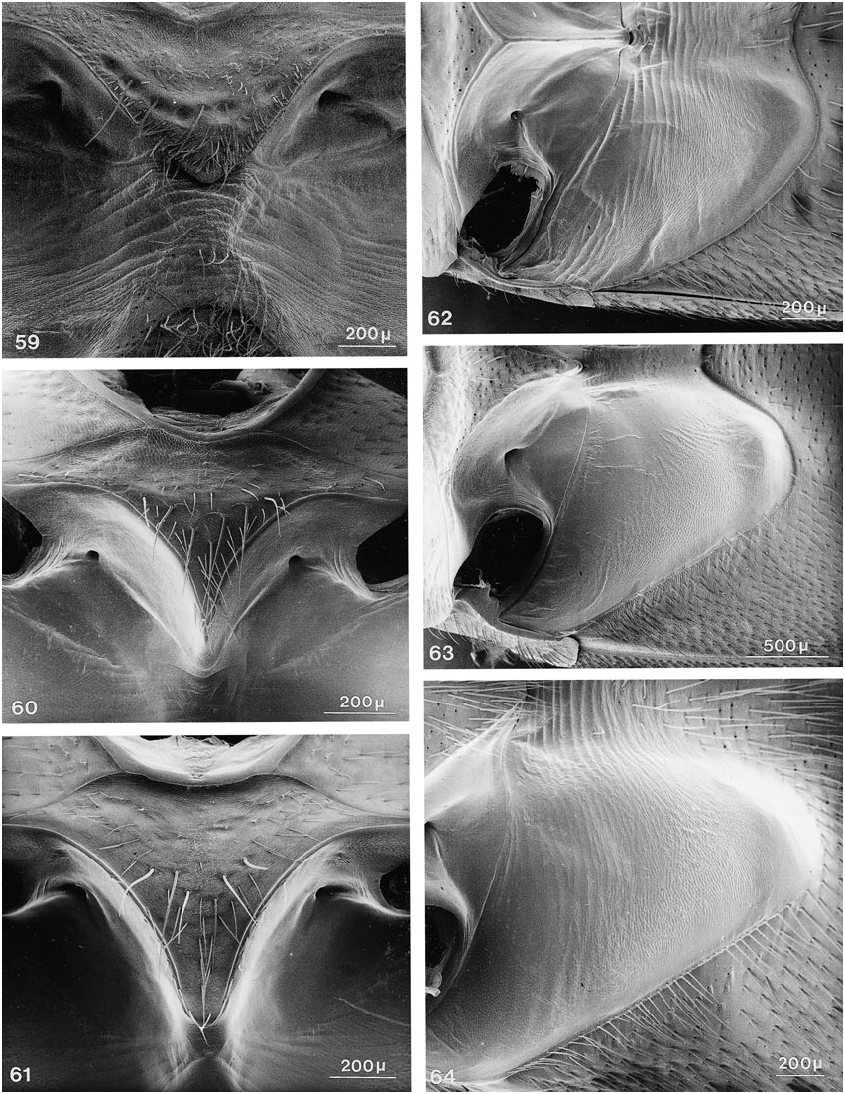
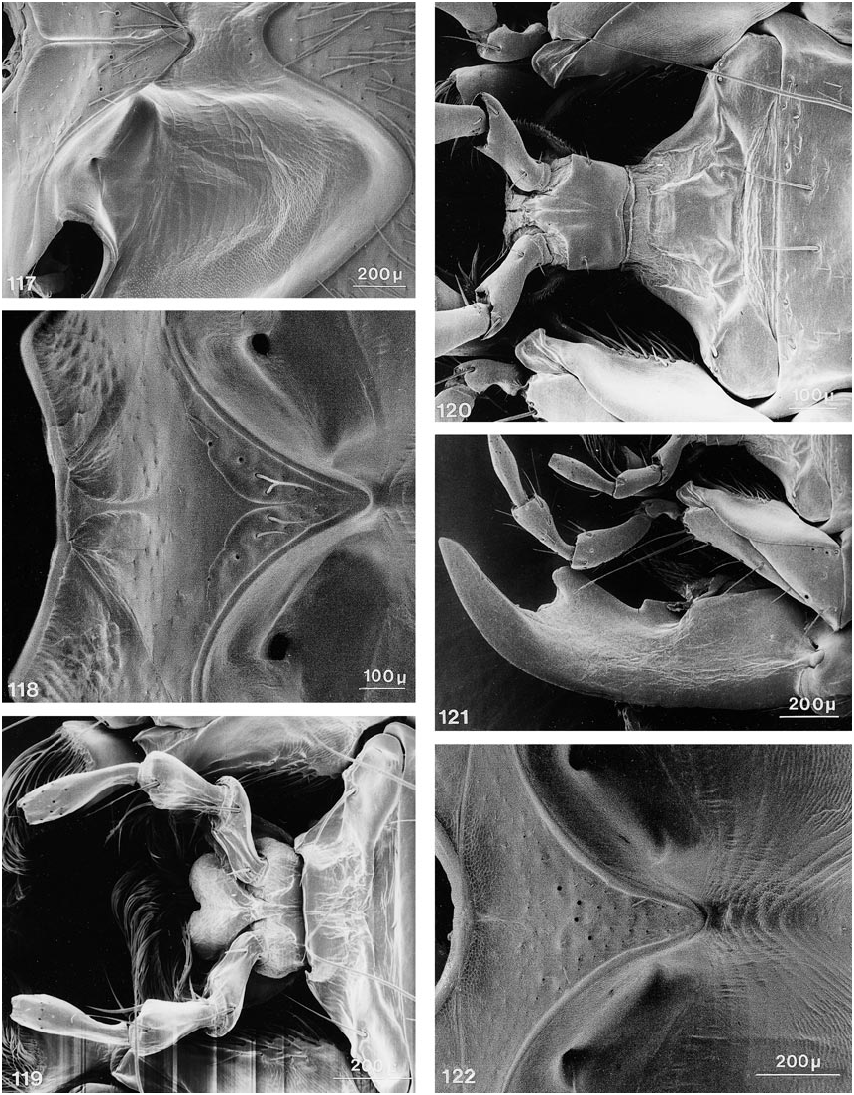
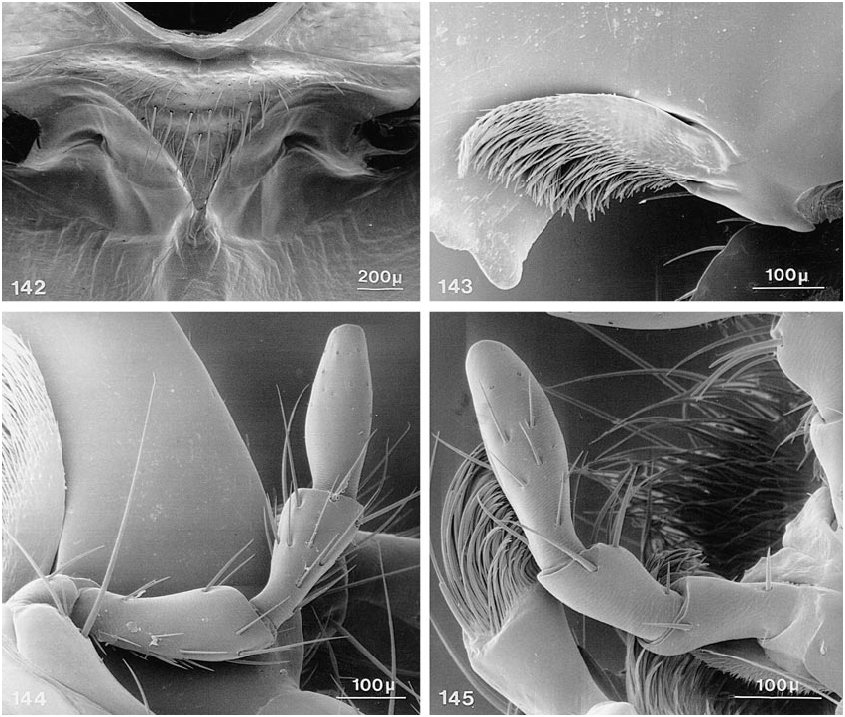
COMMENTS: Members of this genus share the following character states: ( 1) mandibular prostheca bilobed, with basal group of long ciliae and sparser distal group originating on separate supporting arm; and ( 2) long serial setae on mesosternum situated on anterior edges of enlarged, moreorless pitlike punctures, in subtransverse to broadly Ushaped pattern ( fig. 59 View Figs ).
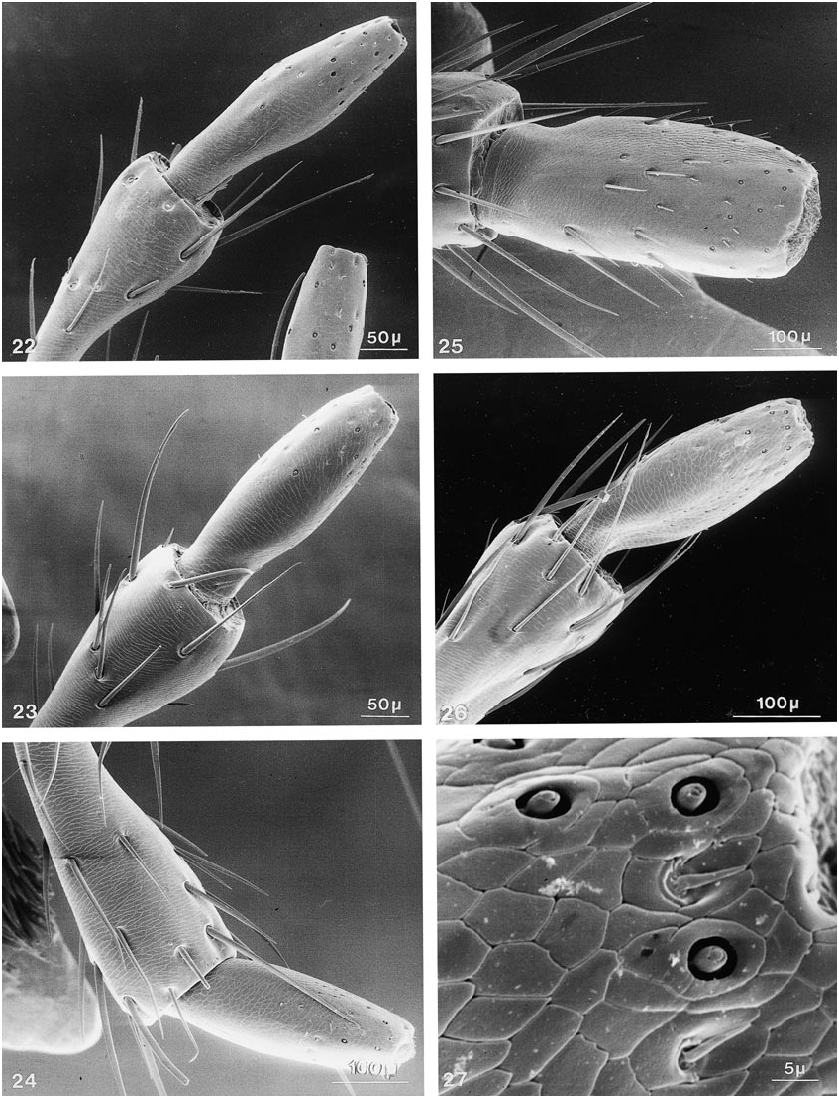
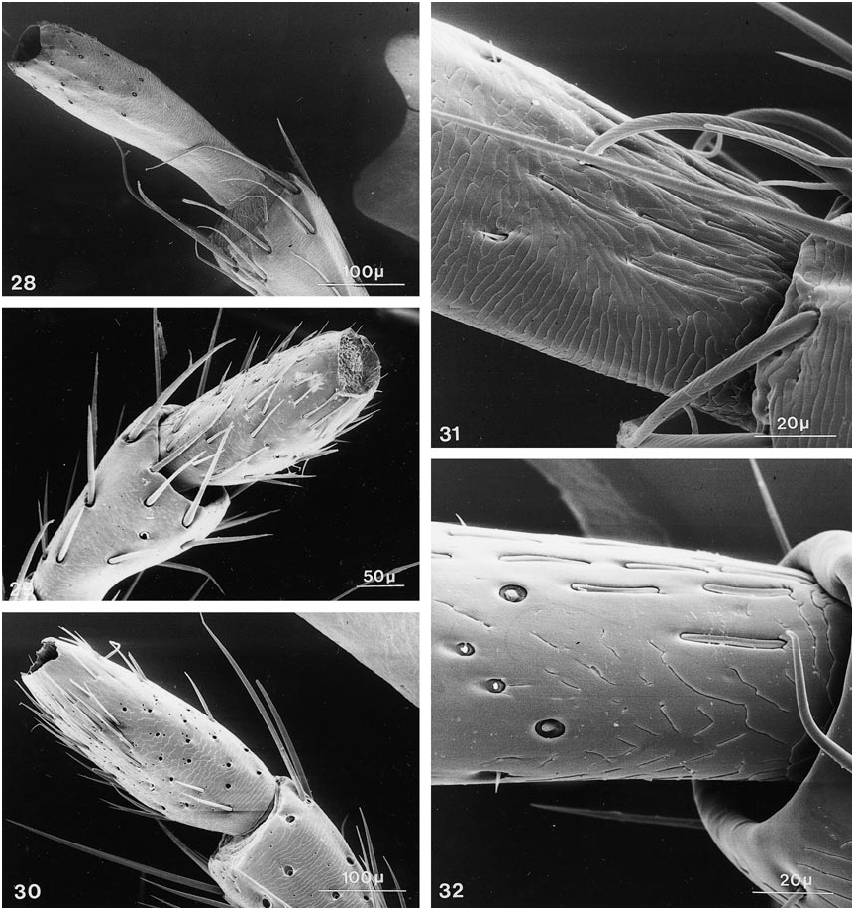
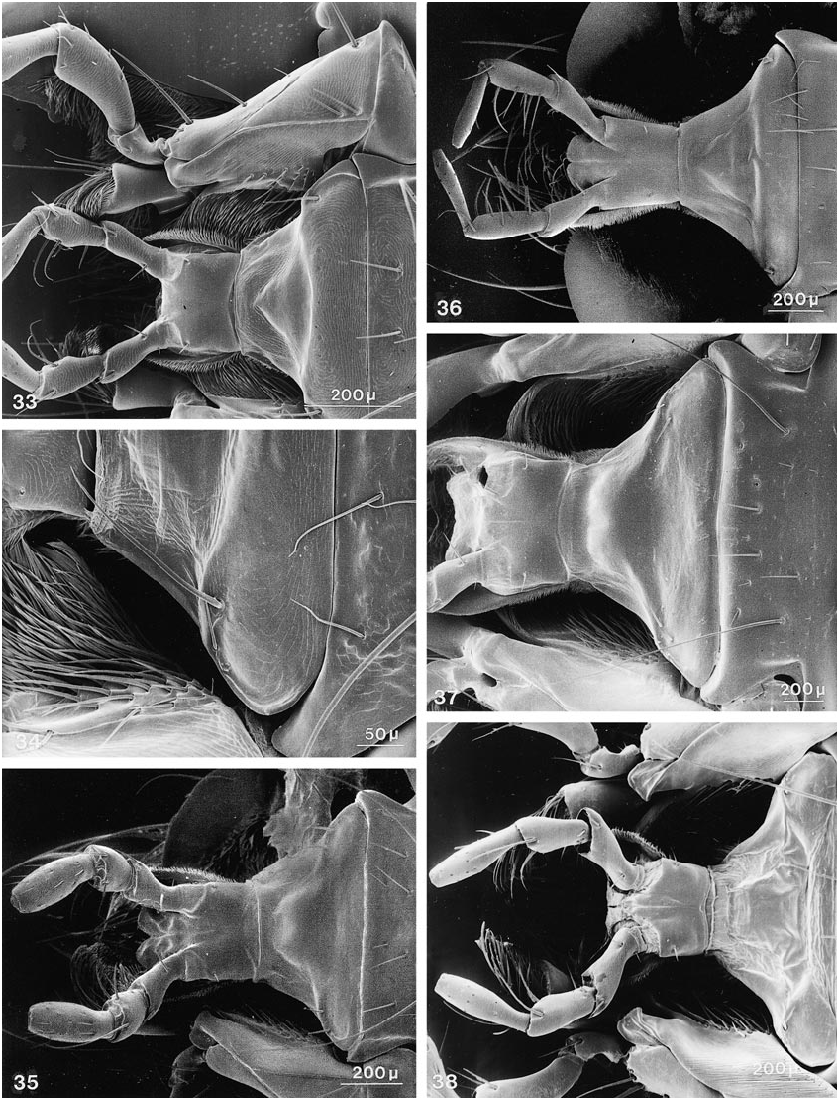
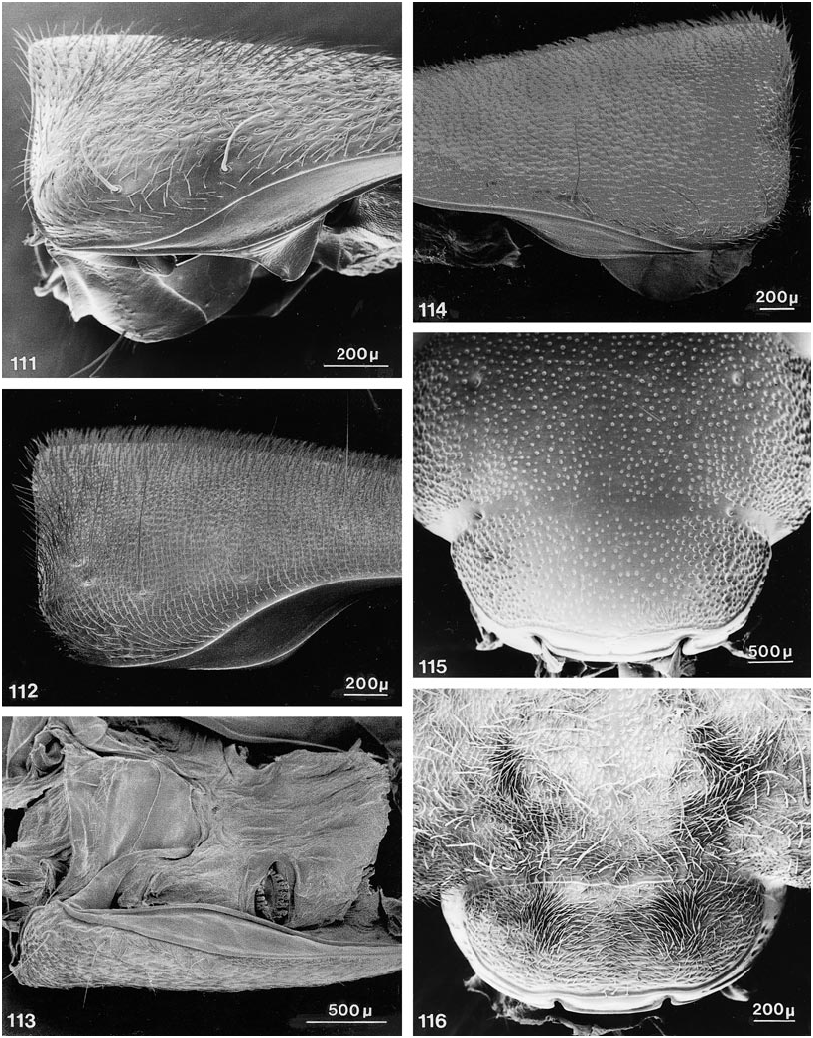
Further character states include: right mandible with one broad tooth at middle, left mandible with two teeth or with one stout, apically emarginate tooth at middle, with all teeth in same (dorsal) plane; maxillary palpus with apical segment moderately elongate, distinctly fusiform, narrower and subequal in length to segment three, asetose ( figs. 23 View Figs , 28 View Figs ); apical segment of labial palpus distinctly fusiform, narrower and markedly longer than segment three, usually with a few, sometimes with many, fine setae ( fig. 38 View Figs ); mentum usually with semicircular fine basal carina extending variably between basolateral angles ( figs. 119, 120 View Figs ); postmandibular ridge rudimentary to extended less than halfway to infraorbital puncture, infraorbital puncture from 4 to 10 diameters distant from margin of eye; postgenal ridge usually broadly obsolete and often deflected obliquely anteriad toward gula, especially in members of Parabemus ; superior line of pronotal hypomeron markedly and relatively abruptly deflected ventrad, before or at most at middle of pronotum, with its anterior portion situated markedly below basal portion in lateral view ( fig. 111 View Figs ), deflected portion of pronotal disc visible in ventral view from behind pos terior margin of epimeron (especially in members of Parabemus ) to near the posterior pronotal angles (especially in Dinothenarus [ Dinothenarus ]); pronotal epimeron ranging from moderately developed to virtually absent; dorsal apicolateral lobe of hind coxa often with more than three (up to six) distinctly spinelike coarser setae.
Subgenus Dinothenarus Thomson, 1858 . Protabemus Scheerpeltz, 1966 ( type species:
Staphylinus xanthocephalus Kraatz, 1859 ).
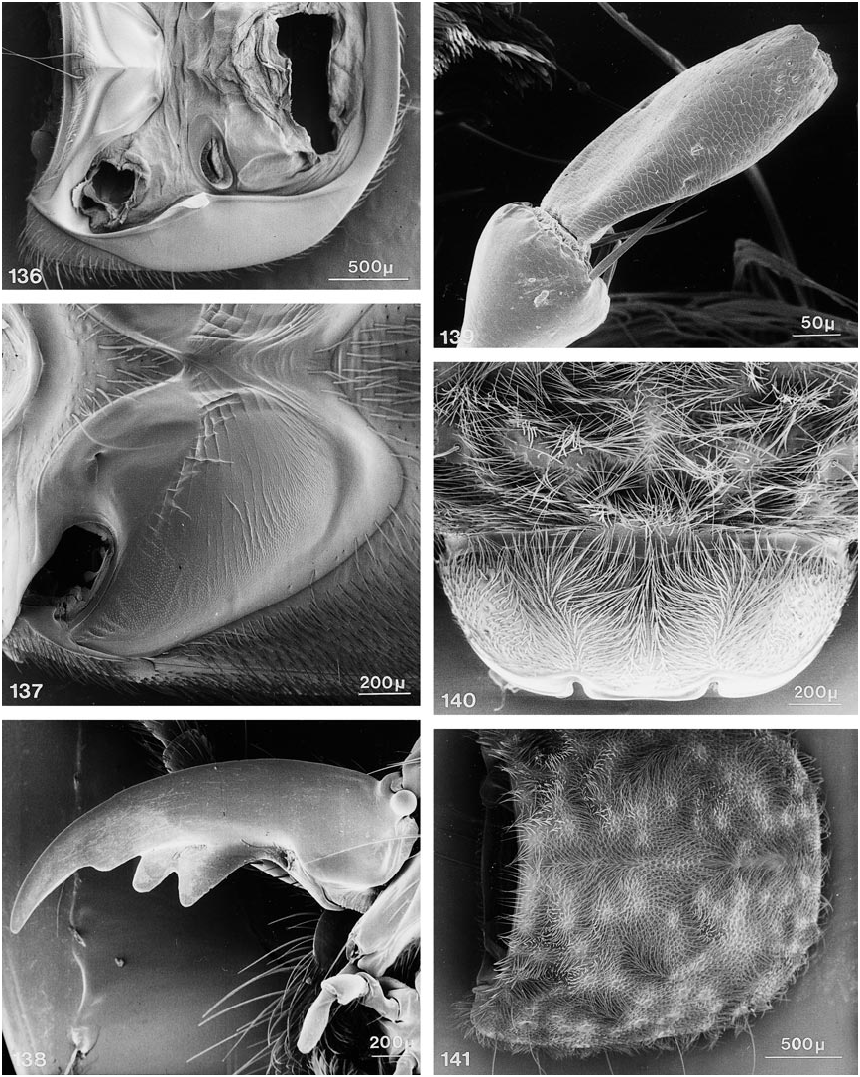
COMMENTS: Members of this subgenus share the following character states: body extensively covered with lush, variegated pubescence of tomentose character ( figs. 140, 141 View Figs ); at least mandibles and adjacent areas, and usually large portions of ventral surface of head bicolored, with distinct testaceous to rufotestaceous areas; ligula virtually entire ( fig. 38 View Figs ), rarely incised apically (e.g., in D. xanthocephalus ); anteromedial portion of prosternum highly prominent, forming a rounded knob which slopes abruptly anteriad and markedly delimits the transverse marginal depression; sternacostal ridge distinctly arched posteriad toward keel, usually coming close to posterior margin of furcasternum ( fig. 54 View Figs ); mesosternal intercoxal process with margins more or less straight, converging subrectangularly until just before rectangular apex, margins without bead apically, slightly explanate; mesocoxal acetabulum only slightly depressed behind projection; margins of anterior metasternal projection never converge at less than right angle, although apex varies from moderately to broadly rounded; disc of metasternum always horizontal, on same plane as body.
The subgenus contains six Palearctic and one Nearctic species at present.
Subgenus Parabemus Reitter, 1909 ( stat. nov.) ( type species: Staphylinus fossor Scopoli, 1772 ).
Parocypus Bernhauer, 1915 ( syn. nov.) ( type species: Staphylinus dehradunensis Bernhauer, 1915 ).
Hypabemus Scheerpeltz, 1966 ( syn. nov.) ( type species: Staphylinus chrysocomus Mannerheim, 1830 ).
COMMENTS: Members of this subgenus share the following character states: body
with patches of variegated pubescence; ventral surface of head piceous to dark rufous, without distinctly bicolored areas; ligula emarginate or bilobed ( fig. 119 View Figs ), anteromedial portion of prosternum usually only moderately prominent, with distinct, but broadly rounded, slope anteriad; sternacostal ridge slightly oblique, relatively straight to moderately arched posteriad toward keel; mesosternal intercoxal process with margins concave, converging distinctly less than rectangularly, with apex acute to narrowly rounded, margins distinctly beaded, at least near apex ( fig. 142 View Figs ); mesocoxal acetabulum slightly depressed to deeply depressed behind projection, with concavity below apex; some species with metasternum short and at an oblique angle to plane of body.
The type species of both Parocypus and Hypabemus share the character states of Parabemus , as they were presented above, and therefore there is no justification for keeping them as separate subgenera.
This is a new, expanded concept of this subgenus that includes 12 east Palearctic/ Oriental species previously assigned to Parocypus , as well as 5 Nearctic species that were previously listed as members of the genus ‘‘ Staphylinus .’’ One of the Nearctic species ( D. pleuralis ) was assigned by Smetana (1965a: 10) to Abemus (as a subgenus of Staphylinus at that time), but Frank (1979: 236) assigned it correctly to Parabemus , based on larval characters. Additional Palearctic and Oriental species will likely be add ed to this subgenus in the future.
REMARKS ON SOME SPECIES OF DINOTHENARUS
1. Dinothenarus fokiensis (Bernhauer, 1933: 32) ( Staphylinus) . Bernhauer (loc. cit.) described the species as a member of the subgenus Parocypus of Staphylinus and compared it to ‘‘ Staph. dehradunensis .’’ Since Parocypus becomes a junior synonym of Parabemus (see above), Bernhauer’s assignment of this species was quite correct. We were able to study the holotype from ‘‘Foochow’’ in the Bernhauer collection in Chicago and another male specimen, compared with the holotype, from the Scheerpeltz collection, Wien, labelled as follows: ‘‘ China Prov. FoKien G. Siemssen.’’ We confirm that Staphylinus fokiensis is a member of the subgenus Parabemus of Dinothenaru s. The above two specimens were compared to the lectotype of Dinothenarus insignis (J. Müller, 1926) (see below) and were found to be conspecific. The name D. fokiensis therefore becomes a junior synonym of D. insignis ( syn. nov.).
2. Dinothenarus fossor (Scopoli, 1772: 109) ( Staphylinus) . Fabricius (1792: 523) described Staphylinus fossor based on specimen(s) of unknown provenience. Erichson (1839a: 377) cited S. fossor Fabricius as a subsequent reference of Staphylinus fossor Scopoli, 1772 . However, Fabricius (loc. cit.) redescribed the species, using the same name S. fossor as had Scopoli (loc. cit.) before him. The name S. fossor Fabricius, 1792 therefore becomes a junior synonym of Staphylinus fossor Scopoli, 1772 ( syn. nov.).
3. Dinothenarus ganglbauerianus (Bernhauer, 1938: 101) ( Staphylinus) . Bernhauer (loc. cit.) described this species as a member of the subgenus Xanthocypus of Staphylinus . Recently, Hayashi (1995: 45) studied the holotype of this species from Kamikochi, Japan (see Hayashi, loc. cit., for the detailed label data), and redescribed it. He considered the species to be a member of the genus Ocypus , very closely related to ‘‘ Ocypus similis ,’’ but at the same time he pointed out some unique characteristics of the species. We were able to study the holotype and concluded, based mainly on thoracic structures, on the shape of the mandibular prostheca, and on the configuration of the aedeagus, that it belongs to the subgenus Parabemus of the genus Dinothenarus and is related to D. insignis . Dinothenarus insignis occurs in southern China ( Fujian) and supposedly also in Japan (see below). However, D. insignis , which is a large and conspicuous species, was apparently never rediscovered in Japan (see Shibata, 1984: 88), which may indicate that one of Müller’s original specimens may have been mislabelled, and that the species does not really occur in Japan. The same actually applies also to D. ganglbauerianus (see Shibata, 1984: 94); we therefore also believe that the holotype of D. ganglbauerianus is likely mislabelled and that this species in fact does not belong to the Japanese fauna. This would confirm the difficulties Hayashi (1995: 45) expressed, based on different grounds, about the reliability of the type locality of D. ganglbauerianus . The aedeagi of D. insignis and D. ganglbauerianus resemble in general configuration those of some members of the genus Ocypus , which contributed to Hayashi’s (1995: 56) conclusion that D. ganglbauerianus was closely related to ‘‘ Ocypus similis ’’ (see above). However, we believe that this similarity in the configuration of the aedeagus is convergent.
4. Dinothenarus insignis (J. Müller, 1926b: 41) ( Staphylinus) . Müller (loc. cit.) described this species as a member of the subgenus Parocypus of the genus Staphylinus , based on two specimens: one from ‘‘ China merid.’’ and one from ‘‘Japonia.’’ The specimen from China is apparently deposited in the Zoologisches Museum der HumboldtUniversität, Berlin (not seen), the other one in the Natural History Museum, London. We were able to study the latter specimen (female), which is labelled as follows: ‘‘ Ocypus n. sp. /Type [round label with red margin]/Typus/ Japan. G. Lewis. 1910 320/ St. (Parocypus) insignis n. sp. Det. J. Müller.’’ The specimen is in fair shape (the right hind tibia and tarsus are missing, and most of the pubescence of the dorsal side of the body is gone, but the remnants permit reconstruction of its original pattern). The specimen is hereby designated as the lectotype of D. insignis ; the label ‘‘ LECTOTYPE Staphylinus insignis J. Müller A. Smetana des. 1999’’ was attached to it.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.