
Campoplex difformis
|
publication ID |
https://doi.org/ 10.5852/ejt.2021.740.1277 |
|
publication LSID |
lsid:zoobank.org:pub:22EEA961-F134-48B3-9655-39C2BB1CC374 |
|
DOI |
https://doi.org/10.5281/zenodo.4640856 |
|
persistent identifier |
https://treatment.plazi.org/id/03844338-451D-2F78-FF3E-FAF3FEBBFB9E |
|
treatment provided by |
Plazi |
|
scientific name |
Campoplex difformis |
| status |
|
Notes on the identification of species of the Campoplex difformis View in CoL group
In his original boxes at the ZSM in Munich, Horstmann arranged the species of Campoplex into ten groups: borealis, continuus, difformis , discrepans, deficiens , faunus, fusciplica, melanostictus, spurius, and tumidulus groups. Of these, only five (borealis, continuus, difformis , discrepans, and melanostictus groups) have been defined by Horstmann in his works (1985, 2000, 2008).
Horstmann (1985) provided a key to European species belonging to the continuus, difformis , discrepans, melanostictus and spurius groups (the last one included in the melanostictus group in Horstmann (1985), but separated in Horstmann’s original boxes), which can be distinguished from the other European species of the genus Campoplex in having the occipital carina in the ventral half turned outwards, meeting the hypostomal carina at a right angle at the base of the mandible ( Fig. 2E–F View Fig ). Even if the demarcation between these groups is often difficult ( Horstmann 1985), species of the difformis group are characterized by the mesopleuron with scattered and shallow punctures on a coriaceous background (i.e., Figs 3C View Fig , 4A–B, 4D View Fig , 5 View Fig , 8B View Fig , 11B View Fig , 15B View Fig ); posterior margins of the female sixth and seventh metasomal tergites only very slightly concave ( Fig. 2D View Fig ); hind tibia yellowish to red-brown, seldom proximally and distally slightly darker, rarely proximally with a light spot (in C. helveticus Horstmann, 1985 and C. hercynicus Horstmann, 1985 ); hind femur red (with the exception of C. helveticus and C. nigricanae Horstmann, 1980 , with the hind femur brown to black); and hind coxa black (except C. canariensis Horstmann, 1980 that has a red hind coxa).
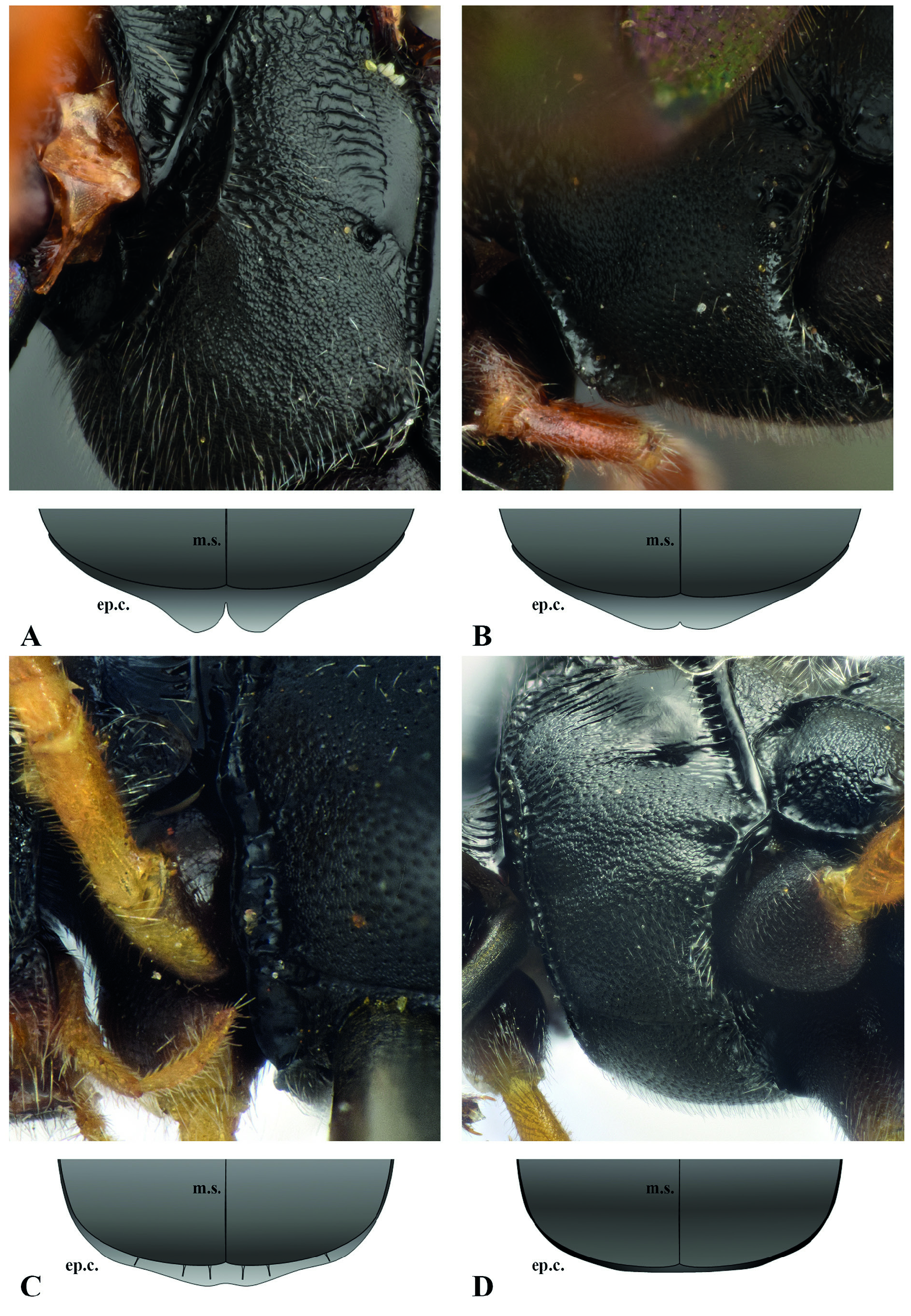
In the difformis group, a few species can easily be recognized by the peculiar shape of the epicnemial carina. In Campoplex melanostoma (Strobl, 1904) (syn. C. anterior Aubert, 1960 ) and C. punctulatus (Szépligeti, 1916) , the epicnemial carina is subventrally abruptly turned towards the ventral hind corner of the pronotum, forming a sharp angled keel ( Fig. 4A–C View Fig ), while it is subventrally more or less straight in the other species of the group ( Fig. 4D–E View Fig ); in C. bilobus (Thomson, 1887) and C. hinziator Aubert, 1980 , the epicnemial carina is ventrally raised and divided into two distinct lobes, thus with a clear notch in the middle separating the two parts ( Fig. 5A View Fig ); in C. hercynicus , the epicnemial carina is strongly raised ventrally, gently rounded and slightly notched in the middle, its width ventrally clearly greater than its width subventrally ( Fig. 5B View Fig ); in C. unicingulatus , the epicnemial carina is evenly raised ventrally and submedially, its width in the middle approximately as high as the width of the fore basitarsus, and not divided in the middle ( Fig. 5C View Fig ).
Females of the remaining species can be separated on the basis of the ovipositor sheath ratio.The ovipositor ratio is less than 1.4 (usually significantly less) in Campoplex tibialis and related species ( Fig. 6A View Fig ), while it is 1.4 or more in species related to C. difformis . In C. restrictor Aubert, 1960 and C. striatus Horstmann, 1985 , the temples are strongly narrowed behind the eyes: imaginary lines connecting the outer side of the eye and temple intersect at the level of the scutellar groove ( Fig. 4F View Fig ), while in species strictly related to C. difformis the temples are not so narrowed: imaginary lines connecting the outer side of the eye and temple intersect at the level of the scutellum or just behind it ( Figs 4G View Fig , 7 View Fig ; with the only possible exception of C. corsicator stat. rev., that has imaginary lines connecting the outer side of the eye and temple intersecting at the level of the scutellar groove or just behind it).
Since the revision of the difformis group by Horstmann (1985), two new species have been described, Campoplex ocellanae Horstmann, 1993 and C. formosanae Horstmann, 2012 , and a third one, C. psilopterus Gravenhorst, 1829 , was recognized as belonging to this group by Horstmann (2000).
According to Horstmann (1993), the identification of Campoplex ocellanae in his key led to C. parvus Horstmann & Yu, 1999 (syn. C. minor Horstmann, 1985 ). Campoplex ocellanae can be inserted at couplet 26 in Horstmann’s key (1985) as follows:
26a.Temples slightly narrowed behind eyes, imaginary lines connecting outer side of eye and temple intersect at the base of the metasoma ( Horstmann 1985: fig. 5). Area superomedia finely coriaceous, not wrinkled; area petiolaris anteriorly coriaceous, posteriorly finely striate; area superomedia and area petiolaris slightly depressed ( Horstmann 1985: fig. 15). Body length about 4 mm. ........................................................................................... C. parvus Horstmann & Yu, 1999 View in CoL
– Temples comparatively more narrowed behind eyes, imaginary lines connecting outer side of eye and temple intersect behind the middle of the mesoscutum ( Horstmann 1985: fig. 6; Horstmann 1993: fig. 4)................................................................................................................................... 26b
26b.Area superomedia and area petiolaris coriaceous and finely wrinkled; area petiolaris in addition with fine transverse wrinkles ( Horstmann 1985: fig. 16). Body length about 6 mm. ................................................................................................ C. sulcatus Horstmann, 1985 View in CoL
– Area superomedia coriaceous and wrinkled only at the lateral margins; area petiolaris entirely and strongly striate, slightly depressed ( Horstmann 1993: fig. 8). Body length about 5 mm. ................................................................................................ C. ocellanae Horstmann, 1993 View in CoL
Campoplex formosanae and C. psilopterus belong to the subgroup of closely related species, together with C. difformis , C. capitator , C. dubitator , and C. unicingulatus , which form a tricky complex of very similar species that are better characterized by their host association ( Horstmann 2012).
Campoplex psilopterus was described from a male by Gravenhorst (1829: 508), probably based on a specimen not completely pigmented (“Suspicor, hoc individuum, coloribus nondum perfecte temperatis, necatum esse”). The species was then cited and redescribed by Ratzeburg (1852: 86), who also described the female and reported a record of Siebold of a male and a female obtained from a species of Psychidae . According to Horstmann (2000), C. psilopterus is near to C. capitator , but it differs in its smaller body size (about 4 mm), slightly narrower face and area petiolaris clearly depressed. With respect to Gravenhorst’s description, Ratzeburg added that the female ovipositor is ¼– 1 / 5 as long as the metasoma. Unfortunately, the original descriptions of Gravenhorst and Ratzeburg and the short note of Horstmann based on the male in Gravenhorst’s collection do not allow this species to be unequivocally characterized. Thus – following Taxapad ( Yu et al. 2016) – C. psilopterus is treated here as species inquirenda.
Campoplex formosanae was reared from the cherry-bark tortrix, Enarmonia formosana (Scopoli, 1763) , in Germany. The species was first treated by authors as C. dubitator (in Tanigoshi & Starý 2003; Jenner et al. 2004, 2005, 2013; Jenner & Kuhlmann 2006; Hunt & Kuhlmann 2007; Hunt et al. 2008; Jenner & Roitberg 2009), while molecular-based studies indicated that it might be conspecific with C. capitator , as molecular differences between the two species were not significant ( Hunt & Kuhlmann 2007; Hunt et al. 2008). However, laboratory tests showed that C. formosanae was unable to develop in Lobesia botrana , the selected host species of C. capitator , and small but constant morphological characters can be found to support C. formosanae as a species distinct from C. capitator and related species ( Hunt et al. 2008; Jenner et al. 2013).
According to Horstmann (1985, 2012), Campoplex formosanae has morphologically intermediate characters between C. dubitator and C. unicingulatus ( Horstmann 2012) . It differs from C. unicingulatus in having ovipositor sheath ratio 1.7–1.8 the ( Fig. 8A View Fig ) (ovipositor ratio 1.4–1.5 in C. unicingulatus ) and the epicnemial carina slightly raised ventrally (at most as high as half the width of the fore basitarsus, Fig. 8B View Fig ) (strongly raised ventrally, about as high as the width of the fore basitarsus in C. unicingulatus ). He reports also that flagellar segments in the apical quarter of C. formosanae are “as long as or slightly shorter than wide” ( Horstmann 2012), but actually flagellar segments in C. formosanae identified by Horstmann himself in NMS seem to be relatively longer than those of C. unicingulatus in Horstmann’s collection ( Fig. 9D–E View Fig ). It differs from C. dubitator in having the area petiolaris clearly depressed and almost entirely covered by transverse wrinkles, including the anterior half ( Figs 8C View Fig , 10D View Fig and Horstmann 2012: fig. 7) (only granulate and with no transverse wrinkles in the anterior half and with fine transverse wrinkles in the posterior half in C. dubitator ; see Figs 10C View Fig , 11C View Fig and Horstmann 1985: fig. 10).
Notes on cocoons of the Campoplex difformis group
Cocoons of the following species have been examined: Campoplex capitator ( Fig. 12A View Fig ), C. dubitator ( Fig. 12B View Fig ), C. formosanae ( Fig. 12C–E View Fig ), C. unicingulatus ( Fig. 12F View Fig ), C. punctulatus ( Fig. 13A–B View Fig ), C. restrictor ( Fig. 13C–D View Fig ), C. sulcatus Horstmann, 1985 ( Fig. 13E View Fig ), and C. melanostoma ( Fig. 13F View Fig ).
Species of Campoplex are solitary koinobiont endoparasitoids, mainly of small moths belonging to families Coleophoridae , Gelechiidae , Pyralidae , Tortricidae , and Yponomeutidae ( Aubert 1983; Horstmann 1980, 1985; Shaw & Aeshlimann 1994; Yu et al. 2016). They preferentially oviposit in larvae and complete their development killing the host as prepupa. Sometimes, when unusual larger hosts are attacked, the hosts are killed before they can reach the prepupal stage; also, a few species kill the host when it has pupated ( Shaw & Aeshlimann 1994; Shaw et al. 2016; Broad et al. 2018); the parasitoid spins its own cocoon inside or outside the host’s remains ( Leong & Oatman 1968; Shaw & Aeshlimann 1994; Athanassov et al. 1998; Shaw et al. 2016); in our samples, at least two species – C. formosanae and C. punctulatus – spin their cocoon both externally to the host’s prepupa remains (that are made by the host's final instar skin) or wait for the host to have pupated and spin the cocoon inside the host’s chrysalis ( Figs 12C–D View Fig , 13A–B View Fig ). Cocoons of Campoplex ( Figs 12–13 View Fig View Fig ) are elongate, sub-cylindrical, with rounded poles; cocoon size is related to adult size, so that male cocoons are generally smaller than those of females; in the examined cocoons, the length is about 3 × (± 0.3) in females and about 2.6 × (± 0.1) in males, the maximum width being measured at the equatorial zone. The colour is quite variable, even within the same species, ranging from pure silky white to very dark brown or blackish, with different shades of colour. The CEB can be present or absent, even when looking at cocoons of the same species; when present, the band can be intense white or dark, or sometimes the cocoon appears bicoloured with two thin external dark bands and a lighter internal band. Thickness and texture are variable too, from very thin and translucent (like in C. capitator ) to very thick and opaque, and from smooth to corrugated surface. The loosely woven outer layer can be reduced or thick, giving the cocoon a woolly appearance and hiding the surface details of the dense middle layer.
In several species of Ichneumonidae and Braconidae there is seasonal dimorphism in the structure and robustness of the cocoon, with the overwintering one thicker, darker and tougher than the summer one ( Shaw & Huddleston 1991; Quicke 2015). The cocoons of Campoplex we examined show an evident dimorphism, even if probably not related to seasonality; most of them are from spring-summer generations, which have not entered diapause. Thus, the cause of observed dimorphism has to be sought in the exploited host and host plant. For example, observing a conspicuous series of cocoons of C. capitator reared in the laboratory on Lobesia botrana collected on Daphne gnidium and on Vitis vinifera L. in Italy, we noticed a constancy in the structure and colour of the cocoons, without evident seasonal variation. However, it cannot be excluded that observed variation in other species is due to the presence of further sibling species that are difficult to separate on a morphological basis. Without a better knowledge of intraspecific variation, it remains extremely difficult to reliably assign specimens developed on different hosts and different places to the same taxon on the base of cocoon features and shapes.












No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |