
Aenictus gracilis Emery
|
publication ID |
https://doi.org/ 10.5281/zenodo.207090 |
|
DOI |
https://doi.org/10.5281/zenodo.5063962 |
|
persistent identifier |
https://treatment.plazi.org/id/038287F4-FFC1-1A09-C8DB-32FC47C6F87A |
|
treatment provided by |
Plazi |
|
scientific name |
Aenictus gracilis Emery |
| status |
|
Aenictus gracilis Emery View in CoL
( Figs. 3–4 View FIGURES 1 – 4 )
Aenictus gracilis Emery, 1893: 187 View in CoL , pl. 8, fig. 1; Wilson, 1964: 463, figs. 56, 57, 89; Bolton, 1995: 59. Aenictus martini Forel, 1901: 473 . Type localities: Malaya, Malacca (Pahang and Perak) and Burma, Moulmein. (Synonymy by Wilson, 1964.)
Aenictus martini var. boelianensis Forel, 1913: 20 . Type locality: Sumatra, Bah Boelian and Bah Soemboe. (Synonymy by Wilson, 1964.)
Types. Aenictus (Typhlatta) gracilis View in CoL . Seven syntype workers on three pins (one on a pin, three on another, three on the other) from Sarawak (MCSN, examined). The single specimen mounted on the first pin is selected as the lectotype, the others as paralectotypes. We also examined two syntypes of A. martini from Malacca and three syntypes of A. martini var. boelianensis (both in MHNG); these specimens well agreed with A. gracilis View in CoL .
Measurements. Worker lectotype and paralectotypes (n =7): TL 3.60–3.80 mm; HL 0.70–0.78 mm; HW 0.63– 0.68 mm; SL 0.52–0.65 mm; ML 1.20–1.30 mm; PL 0.25–0.30 mm; CI 80–90; SI 84–104.
Redescription of worker (lectotype and paralectotypes). Head in full-face view clearly longer than broad, with sides convex and posterior margin almost straight and weakly sinuate; occipital carina distinct. Antenna relatively long; scape almost reaching posterolaterial corner of head; antennal segments II–X each longer than broad; II almost as long as each of III–VI. Frontal carina extending beyond the level of posterior margin of torulus, well developed anteriorly and gradually becoming evanescent posteriorlly. Parafrontal ridge short and bluntly pointed at apex. Masticatory margin of mandible with a large apical tooth followed by a medium-sized subapical tooth and 6–8 denticles; basal margin lacking denticles. Mesosoma slender; promesonotum in profile weakly convex dorsally and sloping gradually to metanotal groove; mesothorax almost cylindrical and demarcated from propodeum by a groove laterally, and by a shallow metanotal groove dorsally. Propodeum in profile with weakly convex dorsal outline; propodeal junction angulate; declivity of propodeum shallowly concave, and encircled with a thin rim; opening of propodeal spiracle clearly circular with its diameter about 1.5–2.0 times as long as diameter of postpetiolar spiracle. Petiole distinctly longer than high, with its dorsal outline weakly convex; subpetiolar process weakly developed, low but often triangular anteriorly, with its ventral outline almost straight, and anteroventral corner angulate. Postpetiole slightly longer than high, with its dorsal outline elevated posteriorlly.
Head including mandible and antennal scape extensively smooth and shiny; basal 1/3 of scape finely sculptured. Pronotum and mesonotum smooth and shiny except for the anteriormost portion of pronotum which is punctate; mesopleuron and propodeum with dense punctures. In addition, propodeum bearing several thin, straight, longitudinal rugae. Petiole with dense punctures, while postpetiole dorsally smooth and shiny, on occasion laterally superficially sculptured. Legs smooth and shiny.
Head and mesosoma dorsally with relatively sparse long standing hairs mixed with sparse short hairs; longest pronotal hair 0.40 mm long. Entire body dark reddish-brown. Typhlatta spot located at occipital corner.
Non-type material examined. THAILAND: W. Thailand, Kanchanaburi Prov., Maeklong Watershed Research Station, 29 XI 2003, Sk. Yamane leg., TH03-SKY-143 ( AMK, SKYC, THNHM); S. Thailand, Ranong Prov., Khlong Naka, 31 XII 2000, W. Jaitrong leg., WJT00-KNK01 ( AMK, SKYC, THNHM); S. Thailand, Satun Prov., Tarutao, Ludu waterfall, 200 m alt., 6 III 2008, N. Noon-anant leg., NW08-TH ( SKYC, THNHM); S. Thailand, Songkhla Prov., Khao Nam Khang, 25 VII 1997, H. Okido leg., TH97-HO-177 ( SKYC, THNHM); S. Thailand, Narathiwat Prov., Tao Dang, peat swamp forest, 7 IX 1998, N. Noon-anant leg., WJT98-NW01 ( SKYC, THNHM). MALAYSIA: Selangor, Ulu Gombak, VII-IX 1992, F. Ito leg., MG355 ( SKYC, THNHM); same loc., 26 VII 1998, F. Ito leg., FI98-190 ( SKYC, THNHM); Sabah, Tawau Hills N.P., 9 VII 1996, K. Eguchi leg. Eg96- BOR-017 ( SKYC, THNHM); same loc., 3 II 1993, T. Kikuta leg ( SKYC); Sabah, Danum Valley, 4 XI 1996, K. Eguchi leg., Eg96-BOR-174 ( SKYC, THNHM); Sabah, Sepilok forest, 23 I 1997, K. Eguchi leg., Eg97-BOR-454 ( SKYC, THNHM); Sabah, Poring, Kinabalu, 450–500 m alt., 21 I 1996, K. Eguchi leg., Eg96-BOR-262 ( SKYC, THNHM); same loc., 26 IV 1997, H. Hirosawa leg. ( SKYC, THNHM); Sabah, Mahua, 13 X 2009, T. Yamasaki leg., BN 09-TY01 ( SKYC, THNHM); Sarawak, Miri, Lambir N.P., 31 XI 1997, Sk. Yamane leg. ( SKYC, THNHM); same loc., 1 I 1998, Sk. Yamane leg. ( SKYC, THNHM); same loc., 16 I 1993, Sk. Yamane leg. ( SKYC, THNHM); same loc., 18 IV 1993, Sk. Yamane leg. ( SKYC, THNHM); same loc., 2 VII 2004, Sk. Yamane leg., SR04-SKY-46 ( SKYC, THNHM); same loc., 16 XII 1993, Sk. Yamane leg. ( SKYC, THNHM); Sarawak, Bako N.P., 21-22 IV 1993, Sk. Yamane leg. ( SKYC, THNHM); Sarawak, Bt. Lanjak, Engkari, 25 IV 1994, K. Het leg. ( SKYC); Sarawak, Mulu, 14 XII 1993, Sk. Yamane leg. ( SKYC). BRUNEI: Tasek Merimbun, 14 VIII 1999, K. Eguchi leg., Eg99-BOR-558 ( SKYC, THNHM). INDONESIA: Sumatra, Sitlung, 1-5 I 1993, F. Ito leg., FI-93-250 ( SKYC, THNHM); Sumatra, Ulu Gadut, 17 VIII 1996 ( SKYC); Sumatra, Mentawai Is., Pulau Sipora, Tuapejat, 25 II 2007, Sk. Yamane leg., SU07-SKY-109 ( SKYC, THNHM); E. Kalimantan, Kutai N.P., 7 VIII 1992, Sk. Yamane leg. ( SKYC, THNHM). PHILIPPINES: Luzon, Los Baños, Mt. Makiling, 12 VII 1997, Sk. Yamane leg. ( SKYC, THNHM); Negros Oriental, near Dumaguete, Valencia, Apolong, 27 XII 1998, Sk. Yamane leg., PH 98-SKY-08 ( SKYC, THNHM).
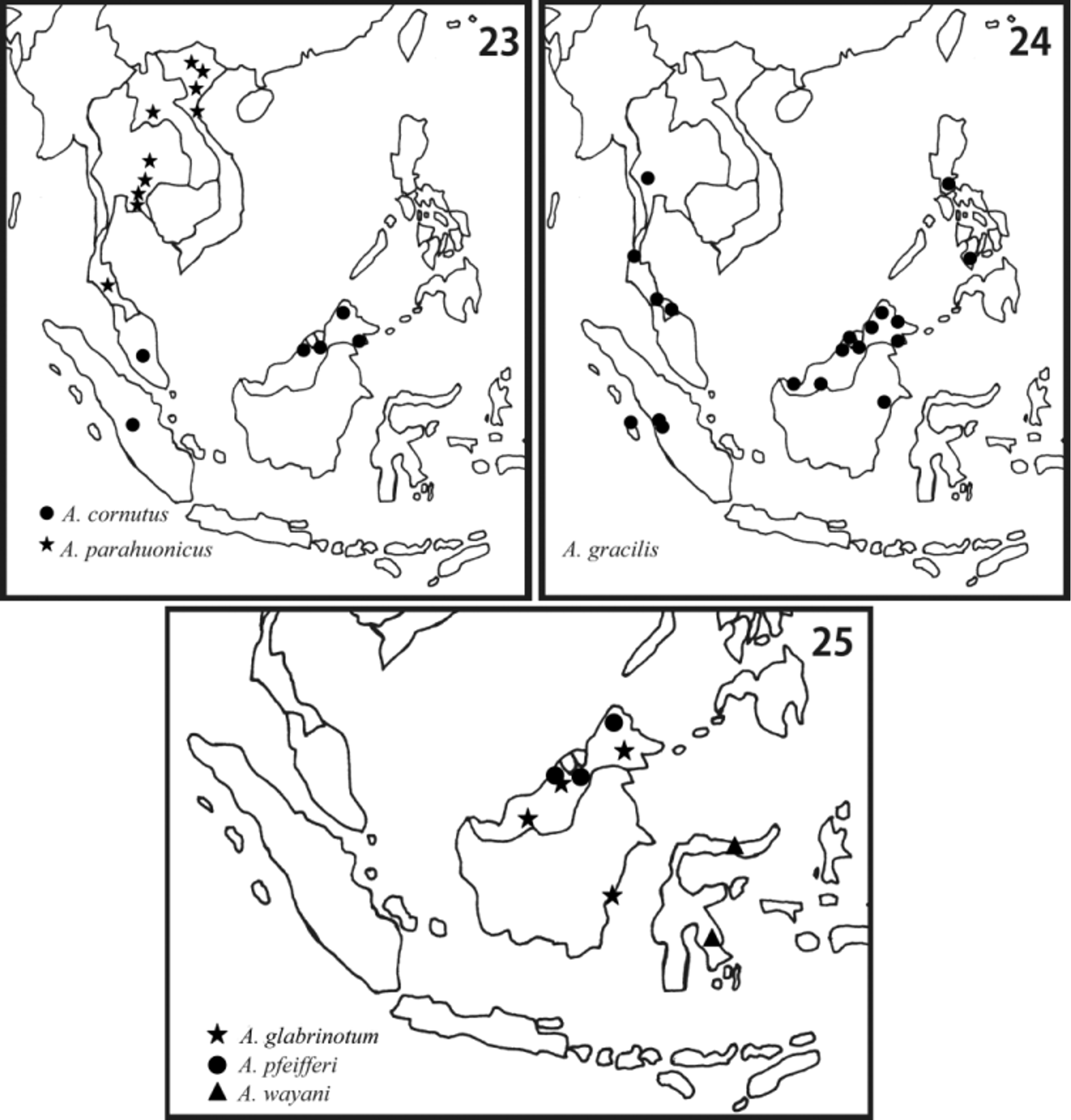
Distribution. Myanmar, Thailand (western part), Malay Peninsula (S. Thailand and W. Malaysia), Sumatra, Borneo (Sabah, Sarawak, Brunei, and E. Kalimantan), and Philippines ( Fig. 24 View FIGURES 23 – 25 ).
Bionomics. A. gracilis is widespread and dominant in the rainforests of Southeast Asia ( Gotwald 1995). The material examined was mainly collected from Sundaland; only two colonies were from the Philippines (Luzon and Nergros islands), and a single colony from western Thailand. Most of these were collected from lowland primary rainforests (less than 500 m alt.). RoŠciszewski and Maschwitz (1994) studied this species in lowland rainforests in Pasoh Forest Reserve, Peninsular Malaysia. Hirosawa et al. (2000) followed many colonies of this species between 600 and 800 m alt. in Sabah, Borneo during his ecological survey on army ants. Schneirla and Reyes (1966) also found many colonies of A. gracilis in open areas around 800 m alt. both day and night in the Philippines. Thus, this species probably ranges from lowland up to 800 m and inhabits both primary and disturbed forests.
Schneirla and Reyes (1966) conducted an ecological study of A. gracilis and A. laeviceps in the Philippines and briefly reported that their food habits are similar to each other, commomnly hunting ant species of the genera Polyrhachis , Camponotus , Crematogaster , and Pheidole , although booty size is generally smaller in A. gracilis . In contrast RoŠciszewski and Maschwitz (1994) found that sympatric species of Aenictus in Pasoh Forest Reserve, Peninsular Malaysia, reduced competition for the same resources by differentially preferring specific taxa, by foraging in different strata or by favoring a particular prey size. The food habits were remarkably different between A. gracilis and A. laeviceps . Their results were supported by Hirosawa et al. (2000), who found A. gracilis forages more frequently on trees and in their canopies than A. laeviceps , which usually forages on the forest floor. Thus, the main prey are arboreal ants in A. gracilis and ground ants in A. laeviceps . Hirosawa et al. (2000) reported that dominant prey genera were Technomyrmex (52.1%), Nylanderia and/or Paraparatrechina (referred to as Paratrechina ) (22.4%) and Crematogaster (11.9%) in the vicinity of Poring, Sabah, Borneo at altitudes of 600– 800 m. J. W. Chapman (cited in Wilson 1964) observed that A. gracilis preyed on other ants such as Anoplolepis gracilipes (referred to as longipes), Camponotus leonardi , Camponotus sp., Crematogaster sp., Leptogenys sp., Paratrechina longicornis , Pheidole sp., Polyrhachis dives , Polyrhachis sp., and also on the social wasp, Ropalidia flavopicta . Chapman (1964) found this species feeding on myriapods, termites, and small staphylinid beetles. RoŠciszewski and Maschwitz (1994) recorded ants of the genera Acropyga , Nylanderia and/or Paraparatrechina (as Paratrechina ), Technomyrmex , and Prenolepis as the prey of A. gracilis .
Remarks. A. gracilis is most similar to A. cornutus in colouration and structure, both sharing the entirely sculptured mesopleuron, metapleuron, propodeum, and petiole. However, this species is distinguished from the latter by the following characteristics: pronotum without horns (pronotum armed with large bilateral horn-like protuberances in A. cornutus ); antennal scape, postpetiole, and legs smooth and shiny (entirely punctate in A. cornutus ); petiole with dense fine punctures (with dense punctures and bearing longitudinal ridges or rugae in A. cornutus ). These two species are sympatric in Borneo, Sumatra and Malay Peninsula.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Aenictinae |
|
Genus |
Aenictus gracilis Emery
| Jaitrong, Weeyawat & Yamane, Seiki 2011 |
Aenictus martini
| Forel 1913: 20 |
Aenictus gracilis
| Bolton 1995: 59 |
| Wilson 1964: 463 |
| Forel 1901: 473 |
| Emery 1893: 187 |