
Microstigmus mirandae, Field, 2023
|
publication ID |
https://doi.org/ 10.1080/00222933.2022.2157345 |
|
publication LSID |
lsid:zoobank.org:pub:E46768B9-FD13-4370-8E31-8D1819B724F4 |
|
DOI |
https://doi.org/10.5281/zenodo.7609005 |
|
persistent identifier |
https://treatment.plazi.org/id/03815C6C-743C-FFD2-E8F3-FD94FEA6FB9B |
|
treatment provided by |
Plazi |
|
scientific name |
Microstigmus mirandae |
| status |
sp. nov. |
Microstigmus mirandae sp. nov.
Recognition
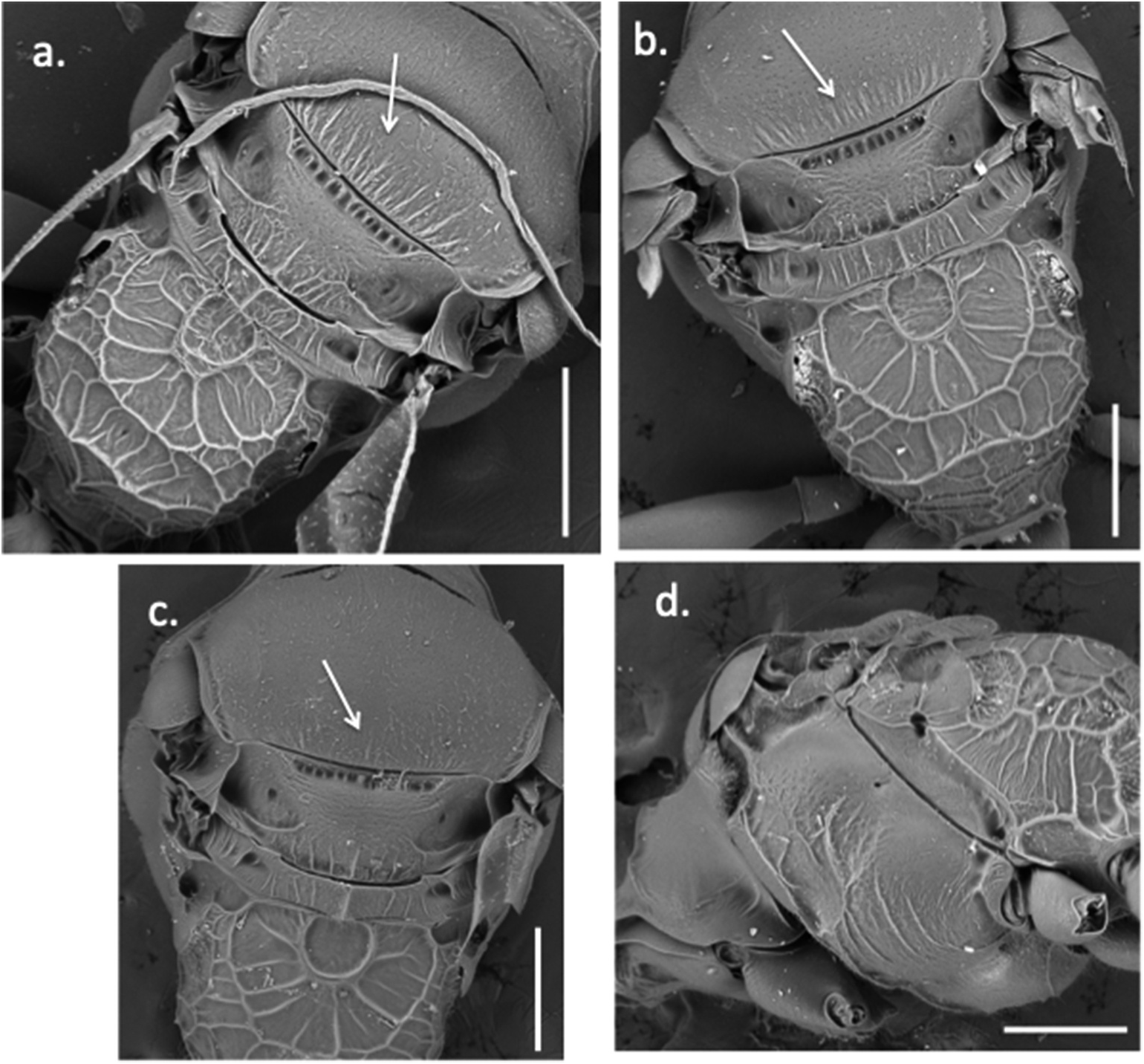
This species has the female 3rd antennal segment (i.e. the first flagellomere) relatively long and thin, like M. lydiae sp. nov. It differs from M. lydiae sp. nov. in: (1) paler overall colouration of head and mesosoma, with the clypeus entirely pale, lacking a thick black line along its top edge ( Figure 1 View Figure 1 (e)); (2) mesopleurae with clear striations latero-ventrally ( Figure 2 View Figure 2 (d)); (3) striations at back of scutum absent or very weak, even at high power ( Figure 2 View Figure 2 (c)); (4) anterior face of male antennal scape white (except yellowish distally at the junction with segment 2), concolourous with lower face and mandibles; (5) male second tergite with a central, narrow, groove-like depression running along the posterior three-quarters or more of the segment.
Description
Female. Length 4 mm. Colouration: head (including underside, face and clypeus; Figure 1 View Figure 1 (e)) and mesosoma predominantly pale orange-yellow; abdomen black ( Figure 1 View Figure 1 (a)). On head, the small area between the ocelli is black in darker specimens. Vertex more or less diffusely brown, with the brown extending down onto face in darker specimens, which also have the underside of the head and mesosoma dark brown/black. In paler specimens, head (including underside and clypeus) pale yellow-orange throughout except vertex diffusely pale brown. Mandibles pale yellowy-white with dark red tips. Antenna pale yellow, more or less brown towards the tips, especially dorsally. On mesosoma, pronotum with anterior dark mark centrally. Scutum, especially at sides, more or less pale brown. Scutellum and metanotum each with central black area, the two areas merged. Mesopleurae pale brown. Underside of mesothorax usually pale or dark brown. Propodeum and abdomen black/dark brown, with a pale mark each side at the base of the second abdominal tergite. Variable pale marks on sternites in some specimens. Coxae and trochanters pale-white, femora partly or entirely (hind femur) dark brown; hind tibiae with a brown mark in some specimens. Legs otherwise pale orange/yellow.
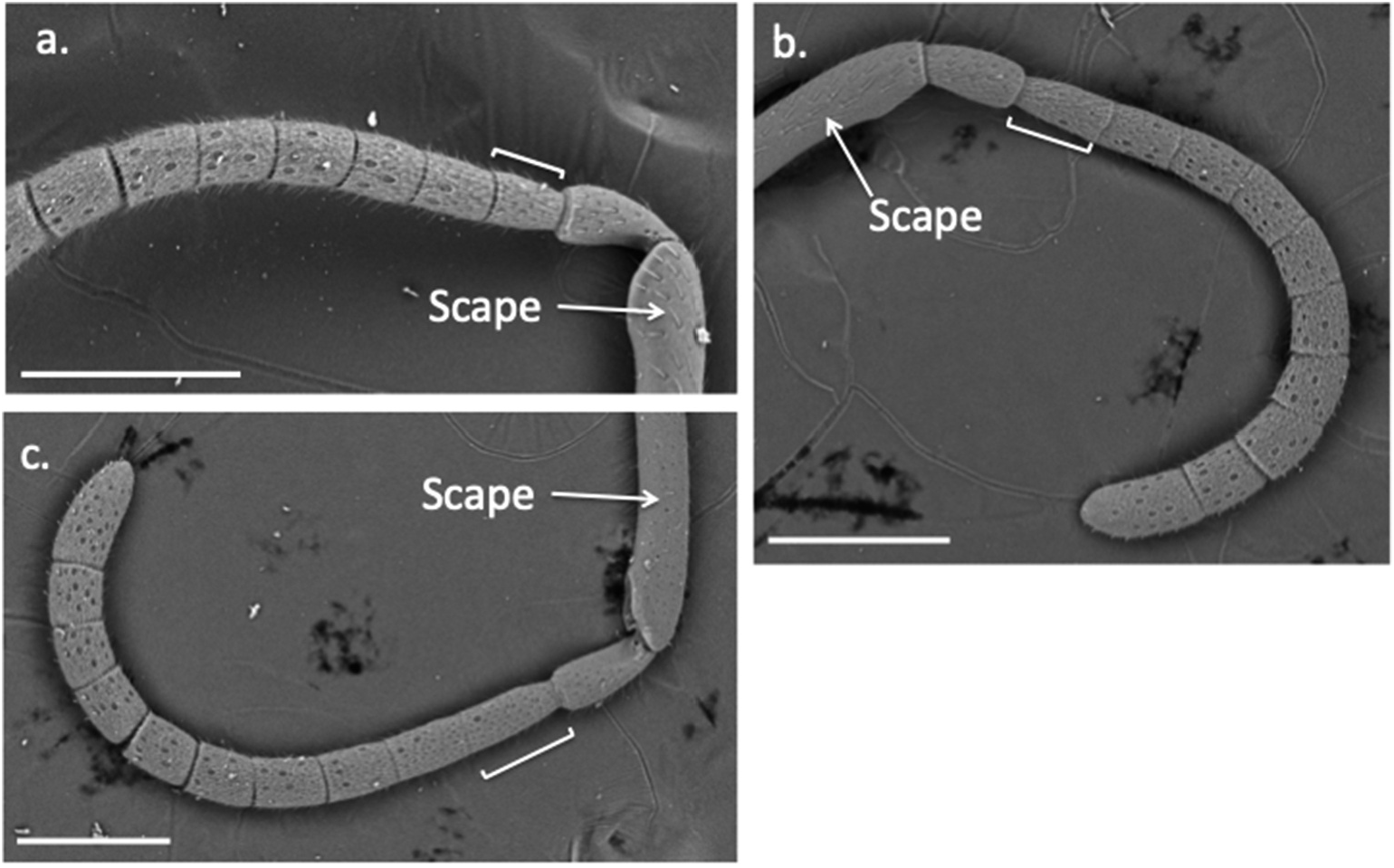
Maximum width of face across the eyes approximately 1.1 times the distance from top of the head to apex of clypeus when head is viewed face-on. Inner margins of compound eyes approximately parallel on face; minimum interorbital distance (across middle of face) approximately 1.1 times interorbital distance near top of the eyes. Ocelli in a not quite equilateral triangle, with the posterior side shorter and the two posterior ocelli separated by slightly less than their diameters. Mandibles with upper tooth shorter than lower tooth. Malar space ( Figure 1 View Figure 1 ) longer than in M. rosae sp. nov., 1.5 times the width of antennal segment 3. Clypeus raised, anterior margin weakly convex. Basal flagellar segments of antennae relatively long and thin: fifth antennal segment 1.6 times longer than wide; 3rd segment 2.5 times longer than wide and, when viewed from above, similar in length to or slightly longer than pedicel ( Figure 3 View Figure 3 (c)). Integument predominantly shining. Frons finely reticulate with sparse, shallow punctures. Scutum shining, impunctate and finely reticulate. A few short striations along the posterior margin of scutum visible on electron micrographs ( Figure 2 View Figure 2 (c)), but these striations short and sparse, not clearly discernable with a binocular microscope at 60 ×. Pronotum with a transverse carina centrally. Scutellum moderately raised centrally. Raised area with a very slight depression centrally and a raised longitudinal keel centrally in the posterior half of the raised area; weaker carinae each side. Metanotum with a sharp, raised longitudinal keel centrally. Mesopleurae with weak sculpturing dorsally, but posteriorly with some some stronger striae latero-ventrally continuing onto the underside of the mesosoma ( Figure 2 View Figure 2 (d)). Omaular sulcus and hypersternaulus well developed, the former wide. Propodeum shining and coarsely sculptured.
Pilosity: compound eye bare, top of head and dorsal mesosoma with dense, short setae visible at high power. In perfect specimens there are two longer, erect setae in the ocellar area, these being approximately the same length as the distance between the centres of the posterior ocelli and less noticeable than in M. lydiae sp. nov. and M. rosae sp. nov. Clypeus and lower edge of mandibles with longer setae. Antennae covered with dense, short setae, especially towards the tips. Dorsal area of propodeum bare, but posterior face with dense setae, these longer than on head and mesosoma. Mesopleurae with short setae, these becoming denser and longer towards the venter and beneath. First two abdominal tergites almost bare, with short setae becoming denser towards the tip of the abdomen, including a few longer setae at the posterior edges of tergites and sternites.
Male. Similar to female except somewhat paler colouration: posterior face and dorsal area of propodeum black or with brown patches but sides of propodeum, scutum, mesopleurae and underside of mesothorax pale yellow/orange. Underside of abdomen largely pale yellow. Antennal flagellar segments are an exception, being almost entirely brown, darker towards the tips. Malar space noticeably longer than in the female ( Figure 1 View Figure 1 ), nearly twice as long as the antennal pedicel. Ventral mesopleural striations less discernible in male. Lower face of male is snowy white, starting half-way down the eyes and including the clypeus and mandibles ( Figure 1 View Figure 1 (f)), except edges of tips of mandibles dark red. Anterior (ventral) side of scape also white. Male posterior tergites bear central depressions. When the abdomen is in its natural position (segments telescoped), these appear as small, groove-like depressions in the centres of tergites 3–7, the overall impression being of a central flattening of the entire posterior dorsal abdomen. In contrast with the male of M. lydiae sp. nov., the surface of tergite 2 also has a central narrow, groove-like depression covering the posterior three-quarters or more of the length of the tergite, this depression being deeper towards the apex of the segment. Tergite 7 is black and concolorous with the other tergites, and has a small, triangular, nick-like emargination at the centre of its posterior margin. Sternite 7 narrowing towards its bluntly rounded, upturned tip.
Type material
Holotype female. ′ ECUADOR: Pichincha, Mashpi Reserve nr Pacto (1050 m). N 00°09.956 ̍ W 078°53.288, 8/12/2016 J.Field ̍. Second label: ′ Nest under butterfly house̍ with ′502̍ written in pencil (NHM). GoogleMaps
Three female and one male paratypes are also deposited at NHM, from the same Mashpi site (all ′ 8/12/2016 J.Field̍, and all with second label: ′Nest under butterfly house̍ and pencil numbers 500, 501, 503 [females] and 500 [male]).
Two further female and one further male paratypes are from the same Mashpi site with similar data but different dates/collectors and no second label (females: ′ 6–7/9/2014 J. Field 32̍ and ′ 1/4/2017 R. Bonifacii MS 1056̍; the first of these two females has the head micropinned separately onto the same piece of plastozoate as the body; male ′ 12/3/2017 R. Bonifacii MS 886̍) . These last three paratypes are currently in the author̍s collection but the intention is to deposit them at NIBEQ once a visit to Ecuador is possible, post-Covid pandemic .
Etymology. This species is named after Miranda Debenham, who has been an inspiration throughout this work.
Biology. Approximately 20 nests of this species were located, all at Mashpi. Nests ( Figure 4 View Figure 4 (c)) were therefore found only at the lower end (600–1086 masl) of the altitudinal range we principally investigated. Nests were found hanging under the leaves of a variety of plants and man-made structures. Nests were typically dark brown in colour with a rough external appearance, apparently constructed from small pieces of plant material (such as wood fragments) interwoven with strands of silk. The nest entrance is located at the apex, where the petiole meets the main nest structure, and the lower part is divided into between 1 and 7 vertically orientated cells, visible externally as bumps on the surface of the nest. Each nest cell contained a single offspring or was empty. Nests were larger (depths typically 13–14 mm, entrance diameter 2.25–2.5 mm) than nests of M. rosae sp. nov. and M. lydiae sp. nov., and had a longer petiole (13–18 mm, mean 15.1) that was always strongly bent at some point between the main nest structure and the substrate ( Figure 4 View Figure 4 (c)). Most nests contained just a single adult female, but there could be up to four females and two males. Offspring were provisioned progressively with leafhopper nymphs ( Cicadellidae ) and no parasites were observed.
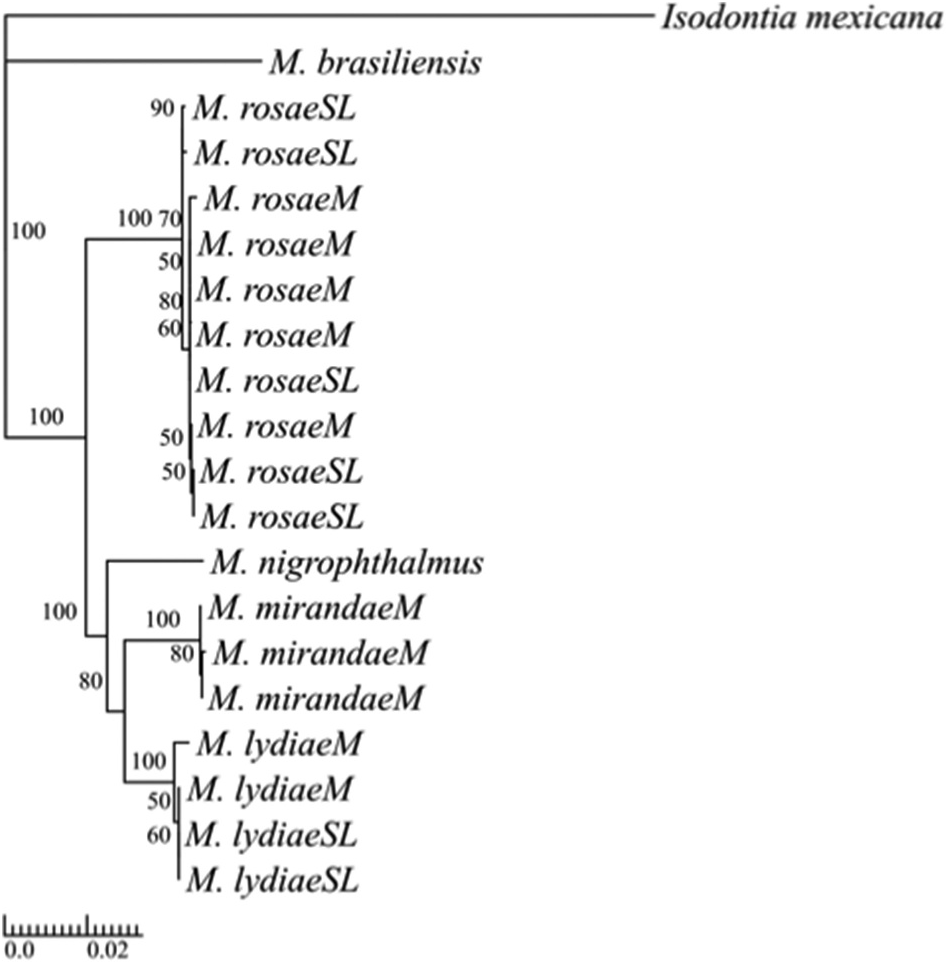
Molecular data. The molecular data were consistent with the species delineations above. First, in the neighbour-joining tree ( Figure 5 View Figure 5 ), individuals of each species identified using the morphological characters above formed a cluster with each other, including individuals from the two main collection sites and across each species̍ observed altitudinal range. Interestingly, the two progressive provisoners that prey on leafhopper nymphs, M. lydiae sp. nov. and M. mirandae sp. nov., clustered together, along with M. nigrophthalmus , the only other progressive provisioner in the tree, which again preys on leafhopper nymphs. Second, the two most similar species, M. lydiae sp. nov. and M. mirandae sp. nov., had alleles in common at only nine of the 29 microsatellite loci scored, at four of which they were both monomorphic for the same allele. Among the remaining 20 loci where there were no alleles common to both species, each species was monomorphic for a different allele at eight loci, while one (n = 10 loci) or both (n = 2) species were polymorphic at the other 12 loci. The microsatellite data will be presented fully elsewhere.


| R |
Departamento de Geologia, Universidad de Chile |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |