
Cyanocharax tipiaia Malabarba & Weitzman, 2003
|
publication ID |
https://doi.org/ 10.5281/zenodo.10813265 |
|
publication LSID |
lsid:zoobank.org:pub:304BA62F-EF75-405C-8FF5-10825986CF0D |
|
DOI |
https://doi.org/10.5281/zenodo.10810795 |
|
persistent identifier |
https://treatment.plazi.org/id/48690CF5-5B6C-4CBB-94B2-E8C867E1C1D1 |
|
taxon LSID |
lsid:zoobank.org:act:48690CF5-5B6C-4CBB-94B2-E8C867E1C1D1 |
|
treatment provided by |
Juliana |
|
scientific name |
Cyanocharax tipiaia Malabarba & Weitzman |
| status |
sp. nov. |
Cyanocharax tipiaia Malabarba & Weitzman View in CoL , new species
(Figs. 1,29-32, Table 4 View Table 4 )
Diagnosis. The low number of perforated scales in the lateral line series (7- 11) distinguishes C. tipiaia from C. alburnus , C. itaimbe and C. dicropotamicus , which have all scales of the lateral line pored. The low number of scales (7- 13) forming a sheath covering unbranched anal-fin rays and anterior 7- 13 branched anal-fin rays differs C. tipiaia from C. lepiclastus , C. macropinna and C. alegretensis (anal-fin base with a sheath of more than 12 small scales covering unbranched anal-fin rays and anterior 12-20 branched anal-fin rays to all anal-fin base).
Description. Morphometric data given in Table 4 View Table 4 . Body elongate and compressed. Dorsal head profile nearly straight to slightly convex. Dorsal body profile from nape to dorsal-fin origin slightly convex, posteroventrally slanted at dorsal-fin base, and nearly straight between dor sal and adipose fins. Ventral body profile convex from lower jaw to pelvic-fin origin, and slightly concave from pelvic to anal-fin origins. Ventral profile along anal-fin base nearly straight to slightly concave. Dorsal and ventral profiles of caudal peduncle nearly straight to slightly concave.
Head small (21.5-23.4% of SL). Eyes relatively large (31.5-37.0% of HL). Maxilla large positioned at an angle of 45 degrees relative to long body axis. Posterior tip of maxilla usually reaching or crossing a vertical line that extends through anterior border of pupil.
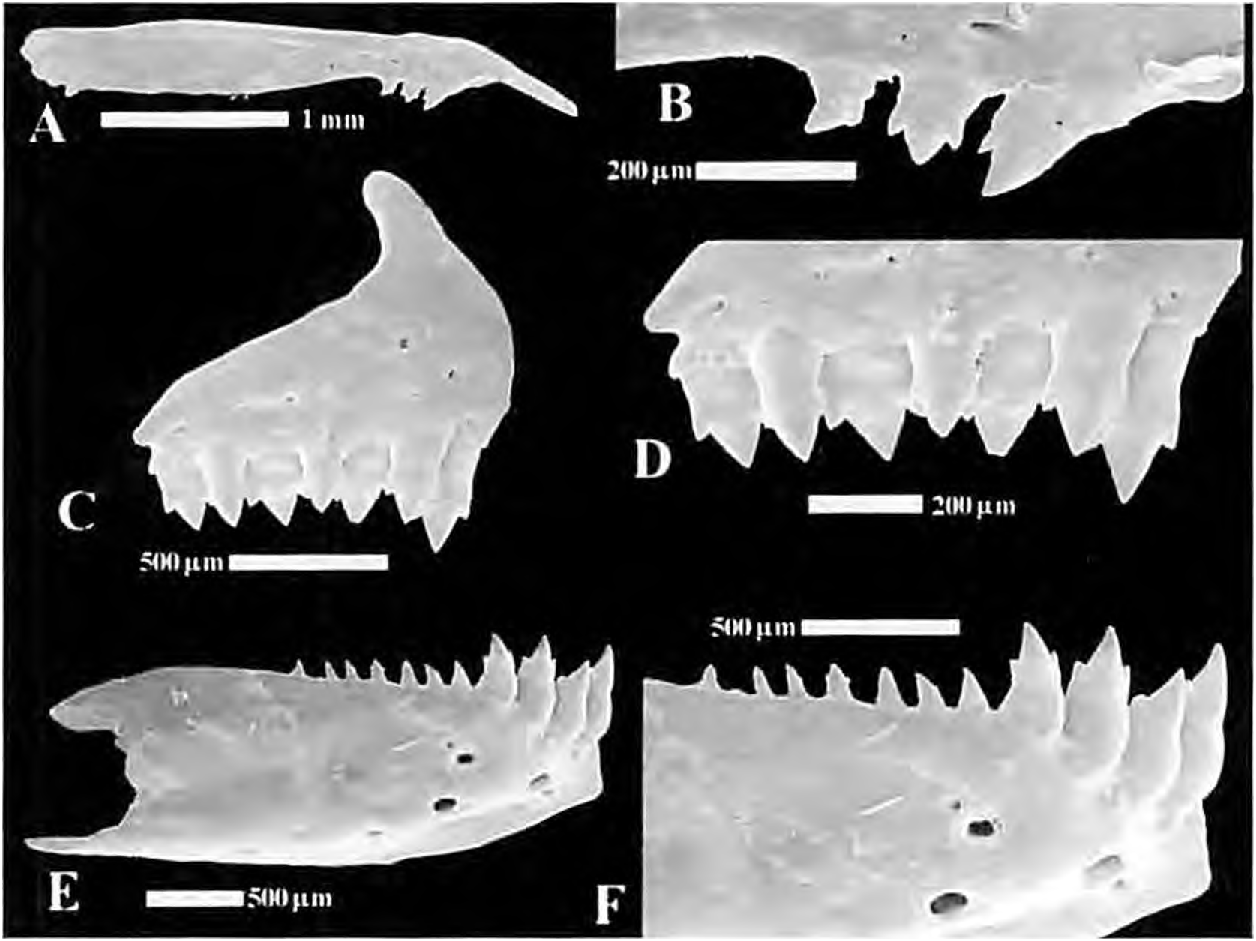
Two series of premaxillary teeth. Teeth of outer series tricuspid, smaller than those of inner series, numbering 3 to 4. Four teeth with 3 cusps in inner series. Two to three maxillary teeth, conical or tricuspid. Four anterior dentary teeth larger with 3 cusps followed by a medium size tooth usually with 3 cusps and a series of 6 to 7 much smaller ones, tricuspid or conical. Second dentary tooth inserted at a lower position in jaws, in such a way that tip of its longest cusp reaches only as high as tip of second largest cusp of first and third dentary teeth ( Fig. 32 View Figure 32 ).
Dorsal-fin rays ii, 8 (n = 15). Dorsal-fin origin near middle of body length. Adipose fin dorsal to posteriormost anal-fin ray insertion.
Anal-fin rays iii-v, 21 -23 (x̄ = 22.3, n = 15). Anal-fin origin ventral to middle dorsal-fin base. Anal-fin distal border slightly concave to nearly straight in all specimens. No mature specimens available to check secondary sexual differences in the anal-fin shape or presence of hooks in mature males.
Pectoral-fin rays i, 9- 10 (x̄ = 9.6, n = 15). Distal end of longest rays extend posterior to pelvic-fin insertion. Pelvic-fin rays i, 6 (x̄= 6.0, n = 15). Pelvic-fin origin anterior to dorsal-fin origin. Distal end of longest rays not reaching anal-fin origin. Mature specimens unavailable to check the presence ofhooks in mature males. Principal caudal rays 10/9.
Scales cycloid. Lateral line incomplete, with 7- 11 (x̄= 8.4, n = 15) perforated scales. Total number of scales in the lateral line series 35-38 (x̄ = 36.8, n = 11). Scale rows between lateral line and dorsal-fin origin, 4- 5 (x̄ = 4.7, n = 15). Scale rows between lateral line and pelvic-fin origin, 2-4 (x̄ = 3.1, n = 15). Predorsal scales, 12- 14 (x̄ = 12.9, n = 15), usually in a regular series. Sheath of scales on anal-fin base consisting of one row with 7- 10 small scales, covering bases of unbranched rays and at least anterior 8 to 12 branched rays.
Vertebrae 36 (n = 13), including Weberian apparatus and posterior half centrum (counts taken from x-ray negatives from MCP 22712 and 22766).
Color in alcohol. Figs. 29-30 View Figure 29 View Figure 30 . Body pale brownish yellow in specimens preserved in formalin long enough to destroy guanine pigment. Lateral body stripe broad and dark gray posteriorly, becoming pale and narrow anterior to vertical through dorsal-fin origin. Humeral spot not discernible. Dark pigmented area present just posterior to upper opening of opercle. Caudal fin darkly pigmented along middle caudal-fin rays to tip of rays. Exposed borders of scales on dorsal portion of body delineated by dark chromatophores. Dorsal and anal fins without distinct marks. Adipose fin slightly dusky. Head black to gray dorsally, especially dark near nape. Sides ofhead whitish yellow.
Sexual dimorphism. In the absence of mature specimens it is impossible to determine whether this species demonstrate the secondary sexual differ ences in the anal-fin shape or the presence of hooks present in congeners.
Etymology. The name tipiaia is a reference to the type-locality, arroio Tipiaia.
Ecological notes. The type locality had moderate water velocity, a rocky bottom, and clear to dark water.
Distribution. Known only from the western small tributaries of the rio Jacuí near Julio de Castilhos, Rio Grande do Sul, Brazil ( Fig. 1 View Figure 1 ).
Holotype: MCP 30471 , arroio Tipiaia , about 13 km North of Julio de Castilhos, on road to Cruz Alta, Rio Grande do Sul, Brazil; 1 Apr 1999.
Paratypes, all from laguna dos Patos drainage , Rio Grande do Sul, Brazil: MCP 22766 (90, 12.2-34.9 mm SL), collected with holotype. MCP 22712 (11: 2 c&s, 34.1 -38.0 mm SL), arroio Felício , about 10 km SSE from Julio de Castilhos; 1 Apr 1999.
Non-type specimens. MCP 26857 (11), same locality as holotype; 28 Nov 2000.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Order |
|
|
Family |
|
|
Genus |