
Berothidae
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3716.2.6 |
|
publication LSID |
lsid:zoobank.org:pub:1D536BCD-4D3B-42B0-98B9-48D8BC31ECE5 |
|
DOI |
https://doi.org/10.5281/zenodo.5677416 |
|
persistent identifier |
https://treatment.plazi.org/id/025F4552-FF99-9517-FF31-FDC7FC37A1B9 |
|
treatment provided by |
Plazi |
|
scientific name |
Berothidae |
| status |
|
Berothidae View in CoL View at ENA indet., larva D
Figs. 8–10 View FIGURE 8 View FIGURE 9 View FIGURE 10
Material examined. Specimen Be 1297, deposited in the amber collection of SF; a third instar larva in a small piece of Baltic amber (precise collecting locality is unknown). The inclusion was bought from the collection of Marius Veta, Lithuania.
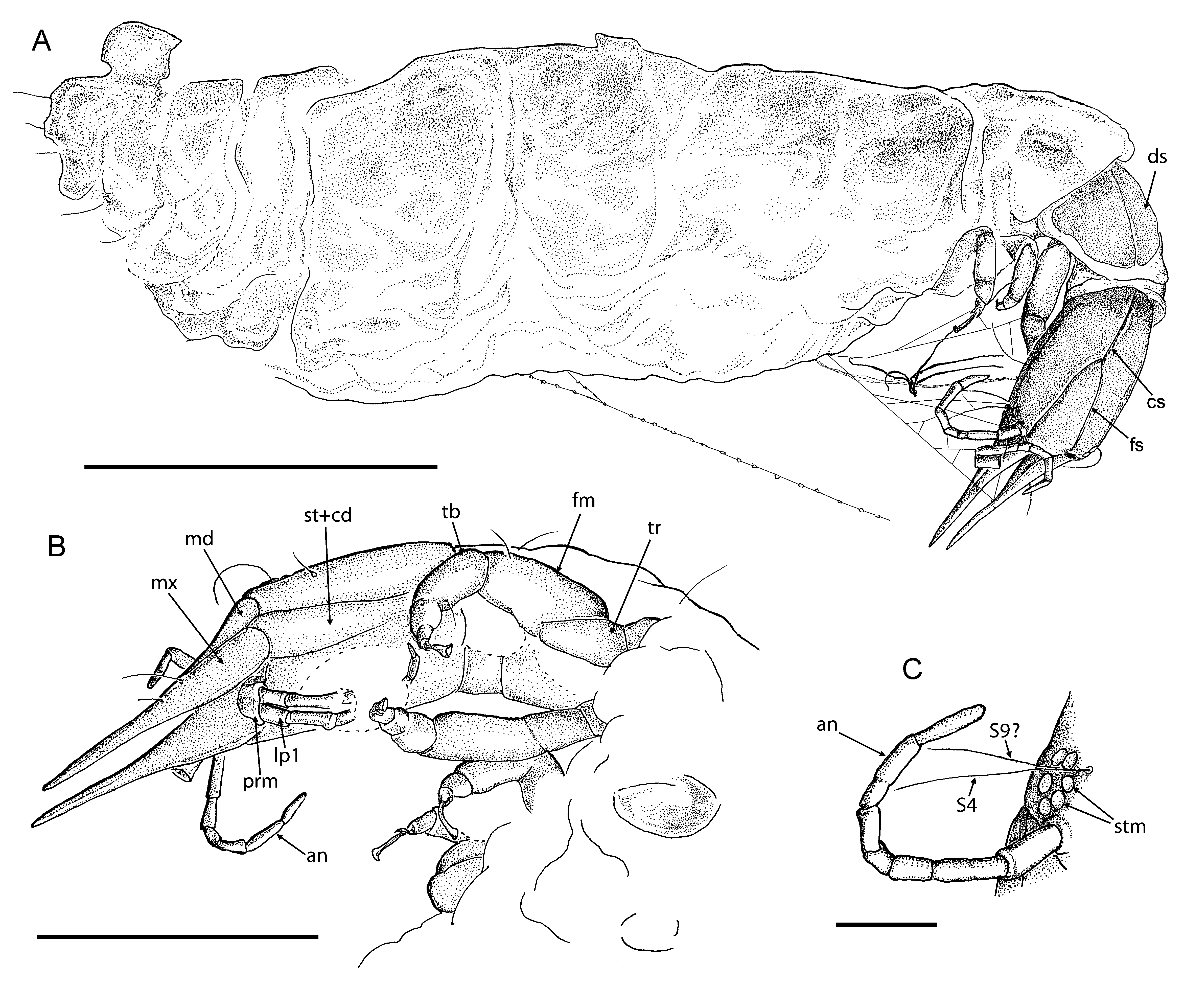
Description. Larva as preserved 3.1 mm long (from tip of cervix to abdomen); actual larval length probably about 6 mm as abdomen is distorted and posterior part with anal sucker is flapped over.
Head capsule elongated, 0.5 mm long, 0.3 mm wide (length/width ratio 1.67).
Six stemmata on each side clearly visible; long setae (about 0.2 mm) ventral to stemmata (probably S9). Dorsally, long seta (about 0.2 mm) near stemmata (S4); sockets of lateral clypeal (trichobothrian) setae on anterior part of capsule; S11 and S12 between arms of frontal suture, probably S1 just posterior to frontal suture, and some other (unidentified) setae in postero-lateral portion of head capsule clearly visible ( Figs. 9 View FIGURE 9 C, 10A, B). Ecdysial cleavage lines very distinct; consisting of frontal and coronal sutures. Two arms of frontal suture extending for approximately half of head capsule length, convergent from inner margin of antennal tubercles until they touch. Lateral sutures absent. Coronal suture long. Ventrally, coxopodites of maxillae (stipes and cardo) rather clearly visible; stipes and cardo relatively narrow; boundary between them not distinct. Labium not clearly visible; prementum appears short (poorly preserved). Ventral setae not visible.
Mandibles 0.44 mm long, relatively broad. Maxillary blades very broad basally, gradually narrowed apically; closely associated with mandible to form mandibulomaxillary stilets, not strongly pointed apically.
Antennal sockets rather flat, without distinct antennal tubercle. Right antenna 0.4 mm long, 7-segmented; left antenna probably 6-segmented; no apical seta visible. Segment lengths of right antenna (from base to top): 0.06 mm, 0.08 mm, 0.04 mm, 0.03 mm, 0.04 mm, 0.08 mm, 0.06 mm. All segments 0.02 mm wide, except for basal segment which is 0.3 mm wide. Left antenna broken twice, but probably complete.
Labial palps poorly visible, about 0.3 mm long; probably 6-segmented; second segment longest and terminal segment very short.
Synchroton radiation computer tomography revealed remains of tissue inside of head ( Fig. 10 View FIGURE 10 C). Oesophagus clearly visible, bifurcated towards mandibulomaxillary stilets.
Cervix probably mainly retracted into prothorax; visible length 0.02 mm dorsally, 0.1 mm laterally.
Prothorax about 0.4 mm long (as preserved); with two large dorsal sclerites that touch each other at midline. Mesothorax and metathorax crumpled, not measurable; at least mesothorax with indistinctly visible paired oval dorsal sclerites.
Legs rather short, robust. Coxa short; trochanter rather large; femur of right foreleg about 0.25 mm long, stout; tibia shorter than femur; tarsus short; pretarsus with two short claws and long, basally strongly narrowed trumpetshaped empodium. Left midleg apparently missing; left hindleg stout (visible with Synchroton radiation computer tomography).
Synchroton radiation computer tomography revealed enigmatic, rounded structure on ventro-lateral side of thorax (dorsal of left hind coxa), ca. 0.15 mm wide, ca. 0.1 mm long; its surface appears very smooth.
Abdomen looks strongly wrinkled; its segmentation vaguely visible. Distal portion of abdomen curved forward; probably with anal sucker.
Remarks. The strongly deformed, wrinkled abdomen, the absence of one leg, and the broken antenna indicate that the larva was already dead and dry when it became embedded in the resin.
Fine web threads are visible around the head connecting mouthparts to the body. This specimen was probably ensnared by a spider as prey in order to catch or store it. This is corroborated by one silky thread connected to the abdomen and leading away rectangularily from it. This thread is equipped with more than 20 tiny adhesive droplets. These are characteristic of some web spiders. Most probably there are also spider threads wrapped all around the abdomen, though these are not clearly visible. But this does not sufficiently explain its deformation.
Apart from spider predation the larva could have been the victim of another predator or some disease before it became spider prey. Its appearance could be the result of decay.
The rounded, oval structure on the ventro-lateral side of the thorax is either a deformed part of the body, or it might be an egg, deposited by a parasite. Further identification of this structure seems not possible.
Review of knowledge on berothid larvae
Ten subfamilies are currently recognized in Berothidae : the Mesozoic Mesithoninae and Paraberothinae, the Rhachiberothinae and Berothinae with fossil and extant species, and the Cyrenoberothinae, Berothimerobiinae, Nyrminae, Protobiellinae, Trichomatinae and Nosybinae which currently include only extant species (Aspöck & Nemeschkal 1998; Monserrat 2006; Makarkin & Kupryjanowicz 2010; Makarkin et al. 2012). Aspöck et al. (2012) consider the Asia Minor genus Nyrma Navás, 1933 and the Chilean Ormiscocerus Blanchard, 1851 as comprising the subfamily Nyrminae. However, the body, legs and wings of Ormiscocerus are much more similar to the other Chilean genus Berothimerobius Monserrat et Deretsky, 1999 (Berothimerobiinae) than to Nyrma (cf. Aspöck 1989; Monserrat & Deretsky 1999; Penny & Winterton 2007). Ormiscocerus and Berothimerobius might be even synonyms. Eggs of Nyrma are stemmed as in most other berothids (R. Dobosz, pers. comm.), whereas eggs of Berothimerobius are sessile ( Monserrat 2006). Therefore, both subfamilies, Nyrminae and Berothimerobiinae, appear to be valid.
The larvae of few species of the following subfamilies of extant berothids are described or illustrated: Rhachiberothinae, Berothimerobiinae, Nyrminae and Berothinae ( Table 1 View TABLE 1 ). The larvae of the presumed cyrenoberothine Manselliberotha neuropterologorum Aspöck et Aspöck, 1988 are known but neither described nor illustrated (Aspöck & Aspöck 1988, p. 9). A larva from Australia not attributed to a taxon is figured by Riek (1970). Judged from its shape and from the relatively small head, it is probably fully-grown and does not belong to the Berothinae because of the position of lateral sutures, which are located distally to the frontal suture (see Riek 1970: Fig. 29.10G, H; New 1991: Fig. 34.10A, B). Such a position of the lateral sutures is similar to that found in Rhachiberothinae, and it is probably plesiomorphic compared with the condition found in Berothinae, in which the lateral sutures originate from the arms of the frontal suture. Three berothid subfamilies occur in Australia: Protobiellinae, Trichomatinae and Berothinae. The figured larva might belong to Protobiellinae, a sister group to the ‘higher’ Berothidae (i.e., Trichomatinae + Nosybinae + Berothinae) (Aspöck & Nemeschkal 1998) judged from its more ‘primitive’ character states (i.e., four-segmented antennae and palpi, position of the lateral sutures).
The most comprehensive and detailed studies on berothid larvae can be found in the unpublished thesis of Möller (2003), which is provided with numerous SEM photomicrographs and drawings of the African species Podallea vasseana , and the dissertation of MacLeod (1964), who examined the head capsule of the American species of Lomamyia Banks, 1905 . The majority of other descriptions are based on the first instar larvae hatched from eggs in the cage (Tillyard 1916; Toschi 1964; Minter 1990; Monserrat 2006). Third instar larvae are known only for two genera of Berothinae ( Podallea and Lomamyia ) (Gurney 1947; Tauber & Tauber 1968; Möller 2003; Möller et al. 2006).
Hitherto, three fossil larvae of Berothidae have been described, all from the Cretaceous: from the Valanginian/ Hauterivian of Jezzine (Jouar Es-Souss), Lebanese amber; from the earliest Cenomanian of Burmese amber (the age is after Shi et al. 2012); and from the Campanian of Grassy Lake, Canadian amber.
The fossil larva described from Lebanese amber is the oldest known berothid larva. Its family affinity as belonging to Berothidae is undoubted, judged from the structure of its mouthparts, ecdysial cleavage lines and antennae (see Whalley 1980: 161, Figs. 9–10 View FIGURE 9 View FIGURE 10 ). The larva is 6–8 mm long (i.e., probably a third instar), very hairy, and possesses a trumpet-shaped empodium; rounded head; five pairs of stemmata; four- or five-segmented labial palps; antennae with an apical seta (but the number of segments is not clear); and “a prominent epicranial suture” [=ecdysial cleavage lines] (Whalley 1980: p. 161). The structure of the latter character is very important; the photo (Whalley 1980: Fig. 10 View FIGURE 10 ) shows that all three sutures of the ecdysial cleavage lines (frontal, coronal and lateral) are probably present. These structures of this larva are most similar to those of the larva of the extant Chilean genus Berothimerobius (although only a first instar larva of the latter species is known and the structure of the ecdysial cleavage lines is unknown, see Monserrat 2006).
The larva from the Burmese amber (about 4 mm long without mouthparts) is of rather typical berothid appearance (Engel & Grimaldi 2008: Figs. 42, 43). It possesses the following principal character states: apparently six small stemmata; palpi with eight palpomeres, with “apical most palpomere slightly broadened, with rounded apex” (however, Fig. 43 shows four palpomeres with short, acute terminal segment); antennae with 15 “articles” (but Fig. 43 shows the number of segments of antennae to be much less, 5–7 segments, with two terminal setae preserved in one antenna). The structure of the ecdysial cleavage lines is unknown.
The newly-hatched larva reported from Canadian amber as belonging to the family Chrysopidae (Engel & Grimaldi 2008: 30, Figs. 12–14) surely belongs to Berothidae . Its size (about 0.6 mm excluding mandibulomaxillary stylets, too small for chrysopid larvae) and the head morphology are typical of Berothidae : the mouthparts are straight, with maxillae which are broad basally and narrowed apically, not curved as in chrysopids; the antennae are 3-segmented, and bear a long strong terminal seta.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.