
Ilanga fulgens ( Dall, 1907 )
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4732.2.1 |
|
publication LSID |
lsid:zoobank.org:pub:F3FE261C-0865-40A7-AAAA-63791DD836A5 |
|
DOI |
https://doi.org/10.5281/zenodo.3664689 |
|
persistent identifier |
https://treatment.plazi.org/id/0078D113-6301-FFDA-FF0B-8ADEFE84FB09 |
|
treatment provided by |
Plazi |
|
scientific name |
Ilanga fulgens ( Dall, 1907 ) |
| status |
|
Ilanga fulgens ( Dall, 1907) View in CoL (new comb.)
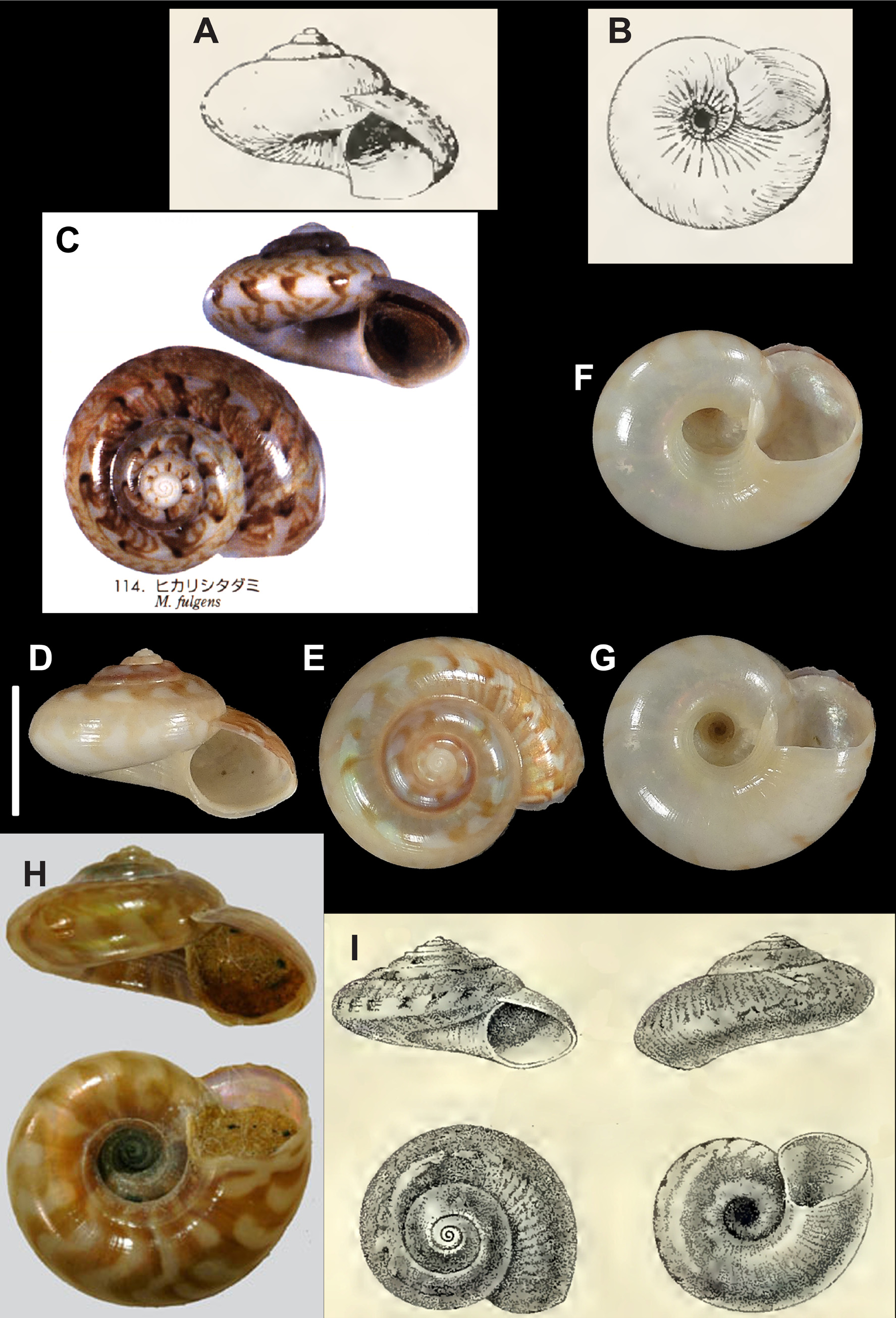
( Figs 11 View FIGURE 11 A–H, 12A-N, 13A–O, 14A-D, Table 5, Key 2: 13)
DNA ref: Ilanga 4 & Ilanga 17 ( Williams et al. 2013; Sumner-Rooney et al. 2016)
COI sequence data: GenBank Accession numbers: Ilanga 4: HF586291 View Materials , HF586292 View Materials . Ilanga 17: HF586260 View Materials , HF586254 View Materials , HF586255 View Materials , HF586256 View Materials , HF586257 View Materials , HF586258 View Materials , HF586259 View Materials , HF586261 View Materials . Ilanga fulgens (this study): MK393381 View Materials .
Note that sequence published as ‘ Microgaza fulgens’ in Williams et al. 2008 is not this species (= Ilanga 19 in Williams et al. 2013).
Microgaza fulgens Dall, 1907: 168–169 View in CoL . Type locality: Sea of Japan, stn 4891, 10°22’.7S, 123°16’.5E, 181 fathoms (390 m) . U. S. Nat. Mus. Cat. No. 110543 .
Microgaza fulgens Kuroda, Habe & Oyama 1971: 42 View in CoL , pl. 12, figs. 15–16.
Microgaza fulgens Kaicher 1986 View in CoL : card TR3-4569.
Microgaza fulgens Higo, Callomon & Goto 1999: 65 View in CoL .
Archiminolia View in CoL fulgens— Marshall, 1999: 24.
Microgaza fulgens Sasaki 2000: 77 View in CoL , pl. 38, fig. 114.
Microgaza fulgens Poppe, Tagaro & Dekker 2006: 129 View in CoL , pl. 68, fig. 3.
Archiminolia fulgens Vilvens 2009: 80–81 View in CoL , figs. 31–34 (part)
Microgaza konos Vilvens 2009: p.88 View in CoL Figs 64–66. Type locality. Taiwan, Bashi channel, TAIWAN 2000, stn DW36, 21°55’N, 120°36’E, 305 m. Syn. nov.
Type material. USNM 110543 ( Dall, 1907) .
Material examined. Northeastern coast of Taiwan. TAIWAN 2001: stn CP75, 24°57’N, 122°02’E, 139 m, 1 dd GoogleMaps . — Stn CP 76, 24°57’N, 122°02’E, 115–170 m, 1 lv (DNA sample MNHN IM-2007-18548) GoogleMaps . —TAIWAN 2002: stn CP162, 22°10’N, 120°38’E, 190–200 m, 2 dd, 1 dd juv 3.7x 7.7 mm GoogleMaps
.
Philippines, Bohol Sea. PANGLAO 2005: stn CP2331, 9°39’N, 123°48’E, 255–268 m, 1 lv (DNA sample MNHN IM-2007-18417), 2 dd juv GoogleMaps . — Stn CP 2332, 9°38’N, 123°44’E, 584–596 m, 2 lv (1 DNA sample MNHN IM-2007-18416), 9 lv GoogleMaps . — Stn CP 2340, 9°29’N, 123°44’E, 271–318 m, 3 lv (DNA samples MNHN IM-2007-18418, 18419, 18420), 19 lv, 1 juv lv GoogleMaps . — Stn CP 2343, 09°27’N, 123°49’E, 273–356 m, 5+15 lv sub, 1+6 dd juv GoogleMaps . — Stn CP 2344, 9°28’N, 123°50’E, 128–142 m, 2 lv (1 DNA sample MNHN IM-2007-18421), 3+1 dd juv GoogleMaps . — Stn DW 2371, 08°35’N, 123°16’E, 172–175 m, 1 dd sub, 3 dd juv GoogleMaps . — Stn CP 2380, 8°41’N, 123°18’E, 150–163 m, 3 lv, 1lv sub, 2 lv juv GoogleMaps . — Stn CP 2381, 8°43’N, 123°19’E, 259–280 m, 1 lv (DNA sample MNHN IM-2007-35122), 1 sub dd GoogleMaps . — Stn CP 2383, 08°45’N, 123°19’E, 338–351 m, 1 dd sub GoogleMaps . — Stn CP 2392, 09°29’N, 123°41’E, 242–400 m, 24 lv, 2 sub lv, 3 juv lv GoogleMaps . — Stn CP 2393, 9°30’N, 123°42’E, 356–396 m, 1 lv (DNA sample MNHN IM-2007-35011), 1 sub dd GoogleMaps . — Stn DW 2402, 09°31’N, 123°42’E, 101–118 m, 2 lv GoogleMaps . — Stn CP 2406, 09°41’N, 123°47’E, 334–389 m, 2 lv GoogleMaps .
South China Sea. ZHONGSHA 2015: stn CP4146, 16°09’N, 114°16’E, 232–314 m, 1 lv (DNA MNHN sample IM-2013-59757) GoogleMaps .
Japan. Wakayama pref., Kushimoto , 90–180 m, 1 lv (C. Vilvens coll.) .
Distribution. Japan, 50–350 m ( Sasaki, 2000); Taiwan, 139–350 m, Philippines, 82–584 m (computed using data of Poppe et al. 2006); Eastern Indonesia, 181–184 m (dd) ( Vilvens, 2009).
Diagnosis. A medium sized Ilanga species with a moderately depressed, conical spire, a subangulate periphery, 4 spiral cords on first teleoconch whorl, weak to very weak subsutural pleats, angulate umbilicus with numerous, sometimes weak, axial pleats all around, bordered by a smooth spiral cord at rim and 6–8 thin spiral cords inside.
Description (supplementary—based on the original description and DNA samples studied). Shell of medium size for genus (H up to 7.7 mm, W to about 12.9 mm), wider than high, shape conical; shell glossy; spire moderately depressed, height 0.53x to 0.66x width, 1.4x–1.8x aperture height; slightly subangulate to almost rounded periphery; umbilicus very broad and deep. Protoconch ca. 200–270 μm wide, 1.25 whorls, rounded, with about ten weak spiral cords sometimes hard to distinguish, only median cord stronger and clearly defined; straight terminal lip, without obvious expansion. Teleoconch up to 4.8 slightly convex whorls, without shoulder, with spiral cords and thin axial threads on first whorls disappearing on next whorls. Suture weakly canaliculated on first whorls. First teleoconch whorl convex, sculptured by 4 thin, evenly spaced, spiral cords appearing immediately; P1 weaker, possibly already disappearing at end of whorl; P2 at edge of angulate subsutural ramp; weak, thin prosocline threads. On second teleoconch whorl, spiral cords disappearing after half whorl, P2 disappearing last; subsutural ramp still present, without spiral cord on rim. On third whorl, axial threads entirely traversing whorl; interspace approximately as wide as thread; subsutural ramp ceasing near end of whorl; no subsutural pleats or possible weak subsutural pleats; narrow translucent (next whorl visible) subsutural spiral band. On last whorls, axial threads very weak but still present; nodules ceasing; few thin abapical spiral grooves around subangular periphery. Aperture broadly triangular; peristome incomplete; outer and inner lip thin, without thickening or weak basal thickening against umbilical rim. Base convex, nearly smooth or with very weak (sometimes hard to see) axial grooves on inner third, reaching umbilical rim. Umbilicus broad (diameter about 30–38% of shell width), central, funnel-shaped, with perspective to apex, with moderately angulate rim marked by a rather weak, thinsmooth spiral cord; moderately convex to almost vertical wall with 6–8 smooth spiral cords and without axial threads inside, spiral cords possibly stronger in adapical part.
Colour: Teleoconch nacreous white or nacreous pink to light brown or orange brown, with variable pattern of brownish regular flames (sometimes in 1 or 2 narrow bands of chevrons), patches or dots; base with regular maculation; protoconch off-white.
Operculum: Corneous, multispiral with central nucleus, light brown.
Remarks. Here, we formally synonymise I. fulgens and Microgaza konos Vilvens, 2009 . The type locality of I. fulgens is Japan, but more recently the name has been applied to samples from the Philippines ( Poppe et al. 2006), whereas I. konos was described for specimens from Taiwan. We synonymise these species based on low levels of genetic divergence and only minor differences in shell morphology. In particular, it appears that the original description of I. konos was based on a subadult sample: compared to the new samples, the holotype of I. konos is smaller with only 4 whorls, a slightly more elevated shape and less numerous axial pleats around the umbilicus, while the other conchological features are similar to those observed in I. fulgens . Umbilical characters were previously the main characters used to distinguish between I. konos and I. fulgens , but with a larger sample size these characters show an overlapping range of sculpture and shapes.
Poppe et al. (2006) also listed Solariella zacalloides Schepman, 1908 ( Fig. 11I View FIGURE 11 ) as a synonym of I. fulgens , but without explanation. We are unable to comment on this opinion because the type material of this species was not studied here.
Molecular data show that specimens from the Philippines identified as I. fulgens cluster together ( Ilanga 17 in Williams et al. 2013), separately to those collected from Japan and Taiwan ( Ilanga 4 in Williams et al. 2013). A single specimen from the South China Sea (new sequence, this study) is distinct from these two groups. Despite small morphological differences among specimens from the different collecting localities (as it can be seen on Figs 11–14 View FIGURE 11 View FIGURE 12 View FIGURE 13 View FIGURE 14 ), we conservatively consider both genetic differences and morphological variation to represent intraspecific variation.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Ilanga fulgens ( Dall, 1907 )
| Vilvens, Claude & Williams, Suzanne T. 2020 |
Archiminolia fulgens Vilvens 2009: 80–81
| Vilvens, C. 2009: 80 |
Microgaza fulgens
| Poppe, G. T. & Tagaro, S. P. & Dekker, H. 2006: 129 |
Microgaza fulgens
| Sasaki, T. 2000: 77 |
Archiminolia
| Marshall, B. A. 1999: 24 |
Microgaza fulgens
| Kuroda, T. & Habe, T. & Oyama, K. 1971: 42 |
Microgaza fulgens
| Dall, W. H. 1907: 168 |