
Filellum sp.
|
publication ID |
https://doi.org/ 10.1080/00222930310001647361 |
|
persistent identifier |
https://treatment.plazi.org/id/002E150C-FFC2-056C-FDE7-FC4F999B4F25 |
|
treatment provided by |
Felipe |
|
scientific name |
Filellum sp. |
| status |
|
Material examined. ANT II- 4 Stn 303, several hydrothecae on Billardia subrufa (RMNH-Coel. 30780); ANT II- 4 Stn 308, several hydrothecae on B. subrufa (RMNH-Coel. 30781); ANT II- 4 Stn 369, numerous hydrothecae on B. subrufa (RMNH-Coel. 30784); ANT II- 4 Stn 450, several hydrothecae on B. subrufa (RMNH- Coel. 30786); ANT II- 4 Stn 460, several hydrothecae on B. subrufa (RMNH-Coel. 30787); ANT V-4 Stn 672, numerous hydrothecae on B. subrufa (RMNH-Coel. 30791); ANT VII- 4 (Epos 3) Stn 281, a few hydrothecae on B. subrufa (MNHN- Hy 1284; RMNH-Coel. 30792); ANT VIII- 5 Stn 16-396, a few hydrothecae on B. subrufa (RMNH-Coel. 30793); ANT VIII- 5 Stn 16-403, several hydrothecae on B. subrufa and Halecium sp. (RMNH-Coel. 30795); ANT VIII- 5 Stn 16-407, a few hydrothecae on B. subrufa and Stegella lobata (RMNH-Coel. 30797); ANT VIII- 5 Stn 16-454, a few hydrothecae on B. subrufa (RMNH-Coel. 30798); ANT VIII- 5 Stn 16-468, several hydrothecae on B. subrufa (RMNH-Coel. 30800); ANT VIII- 5 Stn 16-475, several hydrothecae on B. subrufa and Oswaldella sp. (RMNH-Coel. 30801); ANT VIII- 5 Stn 16-481, several hydrothecae on B. subrufa (RMNH-Coel. 30802); ANT VIII- 5 Stn 16-496, a few hydrothecae on Halecium sp. (RMNH-Coel. 30804);?ANT IX- 3 Stn 207, several hydrothecae on B. subrufa (RMNH-Coel. 30807); ANT IX- 3 Stn 220, a few hydrothecae on B. subrufa (RMNH-Coel. 30808).
Remarks. In Filellum sp. we have included all the infertile material with a hydrotheca similar to that of F. antarcticum and F. serratum . The absence of coppiniae prevents us from identifying it properly.
Ecology and distribution. Filellum sp. comes from depths between 80 and 640 m, on muddy and stony bottoms. It was always found epibiotic on other
hydroids ( Billardia subrufa , Halecium sp. , Oswaldella sp. and Stegella lobata ). The material of Filellum sp. comes from the south and east coasts of the Weddell Sea (off Ronne Ice Shelf, off McDonald Ice Rumples, Caird Coast, off Cape Norvegia and Atka Iceport, Princess Martha Coast, and off Fimbul Ice Shelf, Princess Astrid Coast).
Genus Hebella Allman, 1888 View in CoL
Hebella plana Ritchie, 1907 View in CoL
Hebella striata var. plana Ritchie, 1907: 530 View in CoL , pl. 1 figure 8; 1909: 67; Vanhöffen, 1910: 314; Billard, 1914: 9–10; Jäderholm, 1926: 4.
Hebella plana: Totton, 1930: 156 View in CoL , figure 12a, b; Briggs, 1938: 23; Rees and Thursfield, 1965: 74, 197; Smaldon et al., 1976: 15; Stepan’yants, 1979: 55, pl. 9 figure 9; Boero, 1980: 134; El Beshbeeshy, 1991: 57–59, figure 11; Blanco, 1994a: 161; 1994b: 190; Blanco et al., 1994: 21–23, figures 21, 22; Boero et al., 1997: 13, figure 4 View FIG .
Material examined. ANT II- 4 Stn 372, one hydrotheca on Billardia subrufa ; ANT II- 4 Stn 490, several hydrothecae on Halecium sp. (RMNH-Coel. 30761); ANT II- 4 Stn 502, a few hydrothecae on Stegella lobata (RMNH-Coel. 30762); ANT VIII- 5 Stn 16-403, a few hydrothecae on Halecium sp. (RMNH-Coel. 30763); ANT VIII- 5 Stn 16-407, a few hydrothecae on B. subrufa (RMNH-Coel. 30764); ANT VIII- 5 Stn 16-423, a few hydrothecae on S. lobata (RMNH-Coel. 30765); ANT VIII- 5 Stn 16-475, a few hydrothecae on Symplectoscyphus vanhoeffeni and B. subrufa (RMNH-Coel. 30766).
Remarks. In our material, the length of the hydrothecae is 966–1208 M m, the diameter at the aperture is 225–322 M m and the diameter at the diaphragm 97–145 M m. The length of the pedicel is 325–644 M m and the diameter ca 81 M m.
Totton (1930) raised Ritchie’s variety to specific rank. According to Totton (1930: 156) ‘gonothecae borne singly on hydrorhiza, much larger than hydrothecae, cylindrical, tapering gradually below, with a square, slightly embayed margin provided with three- to five-flapped operculum. The blastostyle bears two, possibly three, medusae which escape from the gonotheca. While still attached the medusa appears to have 4 larger inrolled tentacles and? 4–12 tentacular buds and short manubrium. No pigments spots observed’. Totton remarked that H. plana was the first Antarctic hydroid in which the production of medusae had been observed.
Ecology and distribution. Hebella plana seems to be a shelf species, having been found at depths from 176 ( Billard, 1914) to 550 m ( Jäderholm, 1926) on bottoms of small rocks ( Briggs, 1938) and on muddy bottoms ( Totton, 1930). In our material, it was found between 220 and 722 m depth on muddy and stony bottoms, always on other hydroids ( Billardia subrufa , Halecium sp. , Stegella lobata and Symplectoscyphus vanhoeffeni ). Hebella plana has been found epibiotic on hydroids ( Vanhöffen, 1910; Billard, 1914; Jäderholm, 1926; Totton, 1930; Briggs, 1938; El Beshbeeshy, 1991) and algae ( El Beshbeeshy, 1991). It is also used as substratum by other hydroids (? Opercularella belgicae ). Totton (1930) found gonothecae in January.
Despite the few records available, it seems that H. plana has an Antarctic– Patagonian distribution. Most of the records are from Antarctic waters, both in East and West Antarctica, though there is a record from Patagonian waters by El Beshbeeshy (1991). In East Antarctica it has been collected at 66 ‡ 02’S, 89 ‡ 38’E ( Vanhöffen, 1910) and 66 ‡ 8’S, 94 ‡ 17’E ( Briggs, 1938), both in the Davis Sea, and off Discovery Island ( Jäderholm, 1926) and in McMurdo Sound ( Totton, 1930), both in the Ross Sea. In West Antarctica, H. plana has been found at 74 ‡ 9’S, 22 ‡ W, Coats Coast ( Ritchie, 1907) and from Marguerite Bay ( Billard, 1914). All our material comes from the Weddell Sea (off Ronne Ice Shelf, and off McDonald Ice Rumples, Caird Coast).
Genus Lafoea Lamouroux, 1821
Lafoea dumosa ( Fleming, 1820) View in CoL
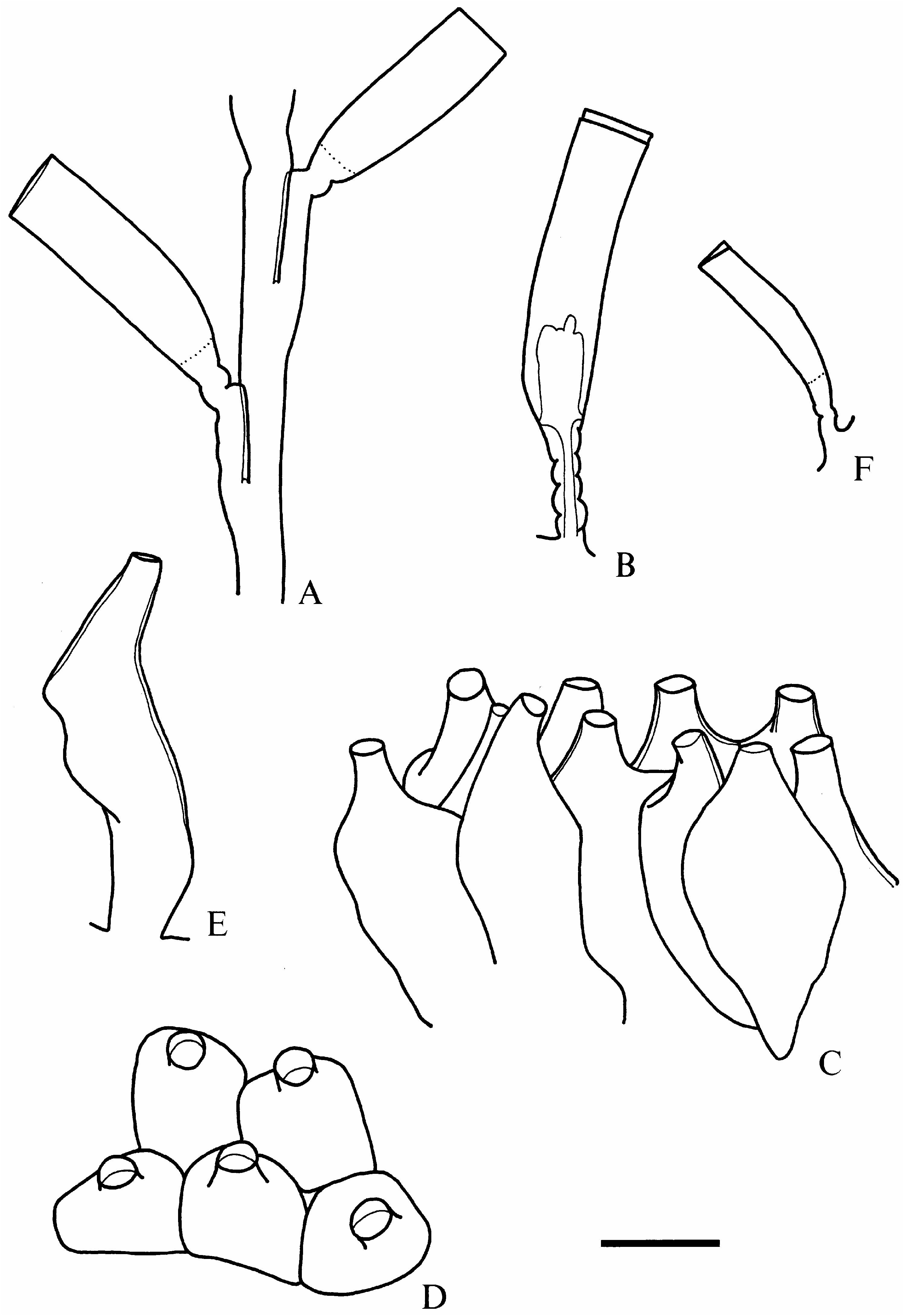
( figure 6F View FIG )
Sertularia dumosa Fleming, 1820: 83 .
Lafoea dumosa: Totton, 1930: 158 View in CoL , figure 14; Millard, 1977: 15; Peña Cantero and García Carrascosa, 1995: 23–25, figure 4A–D View FIG .
Campanularia fruticosa M. Sars, 1851: 138 .
Lafoea fruticosa: G. O. Sars, 1874: 114 View in CoL , figures 16–18; Allman, 1888: 34, pl. 16 figure 2 View FIG ; Totton, 1930: 157, figure 13; Naumov and Stepan’yants, 1962: 76; Vervoort, 1972a: 66, figures 19–21; 1972b: 348, figure 5 View FIG ; Stepan’yants, 1972: 67; 1979: 47, pl. 8, figure 6 View FIG ; Blanco, 1984: 15, pl. 10 figures 22, 23; Blanco et al., 1994: 27–28, figures 27, 28.
Campanularia gracillima Alder, 1856: 361 , pl. 14 figures 5 View FIG , 6 View FIG .
Lafoea gracillima: G. O. Sars, 1874: 115 View in CoL , pl. 4 figures 19–21; Jäderholm, 1903: 273; 1905: 21, pl. 9 figures 2 View FIG , 3 View FIG ; Hartlaub, 1904: 6; 1905: 594, figure P2 View FIG ; Ritchie, 1907: 531; Vanhöffen, 1910: 312; Billard, 1914: 10; Stechow, 1925: 457, figure 24c; Totton, 1930: 158, figure 15; Briggs, 1938: 25.
Lafoea capillaris G. O. Sars, 1874: 115 View in CoL , pl. 4 figures 22–24.
Material examined. ANT I-2 Stn 135, two stems up to 20 mm high on bryozoan, almost without hydrothecae (RMNH-Coel. 30750); ANT VII- 4 (Epos 3) Stn 212, one stem ca 20 mm high (MNHN-Hy 1286; RMNH-Coel. 30751); ANT IX- 3 Stn 220, one colony with a few stolonal hydrothecae and two stems up to 4 mm high, on bryozoans (RMNH-Coel. 30752).
Remarks. In our material, the length of the hydrothecae is 564–660 M m, the diameter at the aperture is 161–177 M m and the diameter at the diaphragm 97–113 M m. The length of the pedicel is 242–290 M m.
Ecology and distribution. In Antarctic waters, L. dumosa has been found from depths between 12 and 520 m ( Stepan’yants, 1979), on stony bottoms ( Peña Cantero and García Carrascosa, 1995); our material comes from 119 to 414 m depth. Lafoea dumosa has been found epibiotic on hydroids ( Totton, 1930; Stepan’yants, 1972; Peña Cantero and García Carrascosa, 1995), but also on ascidians ( Blanco, 1984), bryozoans ( Blanco, 1984; Peña Cantero and García Carrascosa, 1995) and sponges and anthozoans ( Peña Cantero and García Carrascosa, 1995); it was here observed on bryozoans. It has been found with coppinia in May ( Jäderholm, 1905) and December ( Peña Cantero and García Carrascosa, 1995).
Lafoea dumosa View in CoL is considered to have a cosmopolitan distribution. In Antarctic waters, it has been recorded both in East and West Antarctica. In the former, it is known from Gauss Station, 66 ‡ 02’S, 89 ‡ 38’E ( Vanhöffen, 1910), off Oates Land, Cape Adare and McMurdo Sound, in the Ross Sea area ( Totton, 1930), from Commonwealth Bay, King George Land ( Briggs, 1938), from the Davis Sea ( Stepan’yants, 1972, 1979) and from the Friendship Sea, Enderby Land ( Stepan’yants, 1979). In West Antarctica, L. dumosa View in CoL has been found off South Georgia ( Jäderholm, 1905), from Arthur Harbour, Palmer Archipelago ( Vervoort, 1972b), Bransfield Strait, South Shetland Islands ( Stepan’yants, 1979), off Low Island ( Blanco, 1984) and off Shag Rocks, South Georgia and King George Island (South Shetland Islands) ( Peña Cantero and García Carrascosa, 1995). Our material comes from off Elephant Island and the Weddell Sea area (off Riiser-Larsen Ice Shelf and Atka Iceport, Princess Martha Coast). In sub- Antarctic waters, it is known from the Magellan area ( Allman, 1888; Hartlaub, 1904; Naumov and Stepan’yants, 1962; Vervoort, 1972a; Stepan’yants, 1979), the Falkland Islands ( Jäderholm, 1905; Totton, 1930; Vervoort, 1972a), Burdwood Bank ( Jäderholm, 1905; Ritchie, 1907), Kerguélen ( Vanhöffen, 1910; Naumov and Stepan’yants, 1962; Millard, 1977; Stepan’yants, 1979), Crozet ( Vanhöffen, 1910; Stepan’yants, 1979) and Lena Bank ( Stepan’yants, 1979).
Lafoea gaussica Vanhöffen, 1910 View in CoL
( figure 6A–E View FIG )
Lafoea gaussica Vanhöffen, 1910: 312 View in CoL , figure 32; Totton, 1930: 159, figure 16a, b; Stepan’yants, 1979: 48, pl. 8 figure 5 View FIG .
Material examined. ANT II- 4 Stn 450, a few stems up to 8 mm high and several stolonal hydrothecae, on Acryptolaria sp. (RMNH-Coel. 30753); ANT VIII- 5 Stn 16-468, a dozen stems up to 45 mm high growing on tube of benthic organism, with coppinia (RMNH-Coel. 30754); ANT VIII- 5 Stn 16-486, a few stolonal hydrothecae and one incipient stem with two hydrothecae, on stem of stalked sponge (RMNH-Coel. 30755).
Remarks. As Totton (1930) indicated, L. gaussica is easily distinguishable by the large, almost symmetrical hydrothecae, and adnate pedicels ( figure 6A View FIG ). Our material is in complete agreement with the previous descriptions, though we also found a few hydrothecae with completely free pedicels ( figure 6B View FIG ). The length of the hydrothecae is 982–1256 M m and the diameter at aperture is 290–322 M m.
The gonosome of this species was previously unknown. Lafoea gaussica has a characteristic coppinia as also occurs in other species of the genus. The coppinia, ca 15 mm long and 3 mm in diameter, extends on the tube of a benthic organism and on the basal part of two stems. The gonothecae are closely packed ( figure 6C, D View FIG ), being polygonal in dorsal view ( figure 6D View FIG ). The gonotheca is flaskshaped ( figure 6C, E View FIG ) and provided with a long distal neck bearing a circular aperture ( figure 6C–E View FIG ). The coppinia lacks defensive tubes.
Ecology and distribution. Lafoea gaussica seems to be a shelf species. It has been found at depths from 40 ( Stepan’yants, 1979) to 400 m ( Vanhöffen, 1910), on animal debris ( Totton, 1930); our material was collected between 330 and 460 m. We found it epibiotic on other hydroids ( Acryptolaria sp. ), tubes of a benthic organism and the stem of stalked sponges. The fertile colony was collected in February.
Lafoea gaussica View in CoL is an uncommon species, hitherto only known from East Antarctica: from the Davis Sea ( Vanhöffen, 1910), McMurdo Sound (Ross Sea) and off Oates Land ( Totton, 1930), and off Glacier Amery, Friendship Sea ( Stepan’yants, 1979). Nevertheless, our material comes from the Weddell Sea area (off Ronne Ice Shelf, and off McDonald Ice Rumples, Caird Coast), in West Antarctica, therefore indicating a circum-Antarctic distribution of this species.
Family CAMPANULARIIDAE Hincks, 1868 View in CoL
Genus Billardia Totton, 1930 View in CoL
Billardia subrufa ( Jäderholm, 1904) View in CoL
Campanularia subrufa Jäderholm, 1904: 5 ; 1905: 15, pl. 6 figures 4–6 View FIG View FIG View FIG ; Vanhöffen, 1910: 296, figure 16a–c; Nutting, 1915: 42, pl. 5 figures 2 View FIG , 3 View FIG .
Billardia subrufa: Totton, 1930: 151 View in CoL , figure 8b; Briggs, 1938: 18; Broch, 1948: 4; Naumov and Stepan’yants, 1962: 71; 1972: 34, 37; Blanco, 1967a: 252, pl. 1 figures 5 View FIG –8; 1967b: 119; 1984: 17–18, pl. 11 figures 24, 25, pl. 12 figures 26–28; 1994a: 159; 1994b: 191; Stepan’yants, 1972: 67; 1979: 39, pl. 7 figure 4A, B View FIG ; Vervoort, 1972a: 82, figure 25d; Peña Cantero, 1991: 209–213, pl. 6; Bouillon et al., 1995: 78; Peña Cantero and García Carrascosa, 1995: 131–133, figure 60A–E; 1999: 212 et seq.; Genzano and Zamponi, 1997: 290.
Lytoscyphus subrufus: Stechow, 1919b: 81–82 .
Material examined. ANT I-2 Stn 135, two stem fragments up to 40 mm long, with gonothecae (RMNH-Coel. 30685); ANT I-2 Stn 154, one stem ca 80 mm high, with a few incipient gonothecae (RMNH-Coel. 30686); ANT II- 4 Stn 303, one stem ca 140 mm high and a few fragments, with gonothecae (RMNH-Coel. 30687); ANT II- 4 Stn 308, numerous stem fragments up to 100 mm long, with gonothecae (RMNH-Coel. 30688); ANT II- 4 Stn 310, numerous stem fragments up to 120 mm long, with gonothecae (RMNH-Coel. 30689); ANT II- 4 Stn 341, several stem fragments up to 110 mm long, with gonothecae (RMNH-Coel. 30690); ANT II- 4 Stn 369, many stems up to 90 mm high, some on stem of gorgonian, with gonothecae (RMNH-Coel. 30691); ANT II- 4 Stn 372, one stem ca 50 mm high, with remains of a few gonothecae (RMNH-Coel. 30692); ANT II- 4 Stn 386, two stems up to 50 mm high, on axis of gorgonian (RMNH-Coel. 30693); ANT II- 4 Stn 438, numerous stems and fragments up to 180 mm long, with gonothecae (RMNH-Coel. 30694); ANT II- 4 Stn 450, several stem fragments up to 125 mm long, with immature gonothecae (RMNH-Coel. 30695); ANT II- 4 Stn 460, several stems and fragments up to 90 mm high, with gonothecae, on bryozoans (RMNH-Coel. 30696); ANT II- 4 Stn 474, several stems up to 170 mm high and a few incipient stems, with gonothecae, on bryozoans and Oswaldella sp. (RMNH-Coel. 30697); ANT II- 4 Stn 502, several stems up to 120 mm high, with gonothecae (RMNH-Coel. 30698); ANT II- 4 Stn 524, one colony with three stems up to 25 mm high, only gonothecae present (RMNH- Coel. 30699); ANT V-4 Stn 672, several stem fragments up to 70 mm long, with gonothecae (RMNH-Coel. 30700); ANT V-3 Stn 553, one stem ca 120 mm high, with gonothecae but almost without hydrothecae (RMNH-Coel. 30701); ANT VII- 4 (Epos 3) Stn 260, several stems and fragments up to 100 mm long, with gonothecae (MNHN-Hy 1290; RMNH-Coel. 30702); ANT VII- 4 (Epos 3) Stn 261, two detached stems up to 105 mm high and a few smaller stems on unidentified substratum; only a few, badly preserved hydrothecae present (MNHN-Hy 1288; RMNH-Coel. 30703); ANT VII- 4 (Epos 3) Stn 281, one much-fragmented stem (largest fragment ca 200 mm long), with gonothecae (MNHN-Hy 1291; RMNH-Coel. 30704); ANT VII- 4 (Epos 3) Stn 284, three stems up to 40 mm high, on Symplectoscyphus liouvillei ? (MNHN-Hy 1289; RMNH-Coel. 30705); ANT VIII- 5 Stn 16-396, three stems up to 120 mm high, on bryozoans (RMNH-Coel. 30706); ANT VIII- 5 Stn 16-399, several stems up to 180 mm high, with gonothecae (RMNH-Coel. 30707); ANT VIII- 5 Stn 16- 403, numerous stems and fragments up to 180 mm high and smaller stems on polychaete tube and unidentifiable hydroid, with gonothecae (RMNH-Coel. 30708); ANT VIII- 5 Stn 16-405, several stem fragments up to 100 mm long, with gonothecae (RMNH-Coel. 30709); ANT VIII- 5 Stn 16-407, five stems up to 170 mm high, with gonothecae, on bryozoans (RMNH-Coel. 30710); ANT VIII- 5 Stn 16-423, a few stems up to 80 mm high (RMNH-Coel. 30711); ANT VIII- 5 Stn 16-454, several large stems up to 220 mm high, with immature gonothecae (RMNH-Coel. 30712); ANT VIII- 5 Stn 16-456, several stems up to 170 mm high, with gonothecae (RMNH-Coel. 30713); ANT VIII- 5 Stn 16-459, several stems up to 120 mm high, on tube of benthic organism and axis of gorgonian (RMNH-Coel. 30714); ANT VIII- 5 Stn 16-468, a few stems and fragments up to 140 mm high, with gonothecae (RMNH-Coel. 30715); ANT VIII- 5 Stn 16-470, several stems up to 45 mm high, with one gonotheca, on hydrocorals (RMNH- Coel. 30716); ANT VIII- 5 Stn 16-475, several stems up to 210 mm high and smaller stems on polychaete tubes and spine of echinoid, with gonothecae (RMNH-Coel. 30717); ANT VIII- 5 Stn 16-481, three stems up to 70 mm high and a few incipient stems on gorgonian (RMNH-Coel. 30718); ANT VIII- 5 Stn 16-486, four stems up to 140 mm high, and a few stems up to 20 mm high on stalked sponge, with gonothecae (RMNH-Coel. 30719); ANT VIII- 5 Stn 16-492, a few stems up to 300 mm high, with gonothecae (RMNH-Coel. 30720); ANT VIII- 5 Stn 16-496, several stems up to 220 mm high and a few stems on Halecium sp. , with gonothecae (RMNH-Coel. 30721); ANT IX- 3 Stn 123, one stem fragment ca 80 mm long (RMNH-Coel. 30722); ANT IX- 3 Stn 129, one damaged stem ca 90 mm high, with remnants of hydrothecae and gonothecae (RMNH-Coel. 30723); ANT IX- 3 Stn 135, several stems and fragments up to 110 mm high, with gonothecae, on bryozoans (RMNH-Coel. 30724); ANT IX- 3 Stn 158, two stem fragments up to 60 mm high (RMNH-Coel. 30725); ANT IX- 3 Stn 174, three stem fragments up to 70 mm long (RMNH-Coel. 30726); ANT IX- 3 Stn 176, one stem ca 100 mm high, with some incipient gonothecae (RMNH-Coel. 30727); ANT IX- 3 Stn 180, several stems and fragments up to 100 mm high, with one incipient gonotheca (RMNH-Coel. 30728); ANT IX- 3 Stn 207, several stems and fragments up to 70 mm high, on Abietinella operculata , Acryptolaria sp. and bryozoans (RMNH-Coel. 30729); ANT IX- 3 Stn 220, a few stems up to 70 mm high, on bryozoans (RMNH-Coel. 30730).
Remarks. In our material the length of the hydrothecae is 837–1127 M m and the diameter at the aperture is 370–564 M m. The length of the gonothecae is 3140–3542 M m and the maximum diameter is 902–1159 M m.
Ecology and distribution. Billardia subrufa is an eurybathic species. It has been reported at depths from 25 ( Stepan’yants, 1972) to 604 m ( Vervoort, 1972a) on muddy ( Totton, 1930), sandy ( Naumov and Stepan’yants, 1972) and stony bottoms ( Peña Cantero and García Carrascosa, 1995), and on bottoms of rocks and mud ( Peña Cantero and García Carrascosa, 1995), bottoms with a small amount of ooze and with small rocks ( Briggs, 1938), and bottoms of gravels with sponges, hydroids, bryozoans and alcyonarians ( Naumov and Stepan’yants, 1972). Our material comes from depths between 80 and 1030 m, considerably extending its bathymetrical distribution range, on muddy, stony and rocky bottoms and others constituted of fragments of Bryozoa. Billardia subrufa has been reported growing on worm tubes and other organisms ( Totton, 1930), and on hydroids ( Naumov and Stepan’yants, 1972; Peña Cantero and García Carrascosa, 1995). We found it epibiotic on a wide range of substrata: tubes of polychaetes and other benthic organisms, stems of gorgonians, spines of echinoids, stalked sponges, bryozoans and hydroids ( Abietinella operculata , Acryptolaria sp. , Halecium sp. , Oswaldella sp. , Symplectoscyphus liouvillei and hydrocorals). It is used in turn as substratum by numerous species of hydroids ( Campanularia sp. , Filellum antarcticum , Filellum magnificum sp. nov., Filellum sp. , Halecium sp. , Hebella plana ,? Opercularella belgicae , Stegella lobata , Symplectoscyphus sp. ). Fertile colonies of Billardia subrufa have been recorded in January ( Totton, 1930; Briggs, 1938), February ( Peña Cantero and García Carrascosa, 1995), April ( Jäderholm, 1904; Totton, 1930), May ( Vanhöffen, 1910) and December ( Jäderholm, 1904; Briggs, 1938; Peña Cantero and García Carrascosa, 1995); in our material the colonies with gonothecae were collected in October, December, January and February.
Billardia subrufa View in CoL is known from both East and West Antarctica. In the former it has been reported from McMurdo Sound, in the Ross Sea ( Totton, 1930), from Commonwealth Bay, King George Land, and at 66 ‡ 32’S, 141 ‡ 37’E and 66 ‡ 8’S, 94 ‡ 17’E ( Briggs, 1938), off Sabrina Coast, Princess Ragnhild Coast and Queen Mary Coast ( Naumov and Stepan’yants, 1962), off Adélie Land ( Naumov and Stepan’yants, 1972), and from the Davis Sea ( Vanhöffen, 1910; Stepan’yants, 1972, 1979) and Cosmonauts Sea ( Stepan’yants, 1979). In West Antarctica, it has been recorded from the Weddell Sea ( Broch, 1948; Blanco, 1967a), Bransfield Strait, Graham Land ( Vervoort, 1972a), off Shag Rocks, South Georgia, and off Cape Seymour, Snow Hill and Joinville Islands, Graham Land ( Jäderholm, 1904), off Low Island ( Blanco, 1984), at 64 ‡ 10’S, 62 ‡ 40’W and 65 ‡ 11’S, 64 ‡ 10’W (Palmer Archipelago) (Blanco, 1994), and off Powell Island (South Orkney Islands) and King George Island (South Shetland Islands) ( Peña Cantero and García Carrascosa, 1995). In our material, B. subrufa View in CoL was found along the west, south and east coasts of the Weddell Sea (off Larsen Ice Shelf, Antarctic Peninsula, off Ronne Ice Shelf, off Riiser-Larsen Ice Shelf, Cape Norvegia, Lyddan Island and Atka Iceport, Princess Martha Coast, off McDonald Ice Rumples, Caird Coast, and off Fimbul Ice Shelf, Princess Astrid Coast). Billardia subrufa View in CoL has also been reported from the subantarctic Falkland Islands ( Totton, 1930), indicating that it may have an Antarctic–Patagonian distribution.
Genus Campanularia Lamarck, 1816 View in CoL
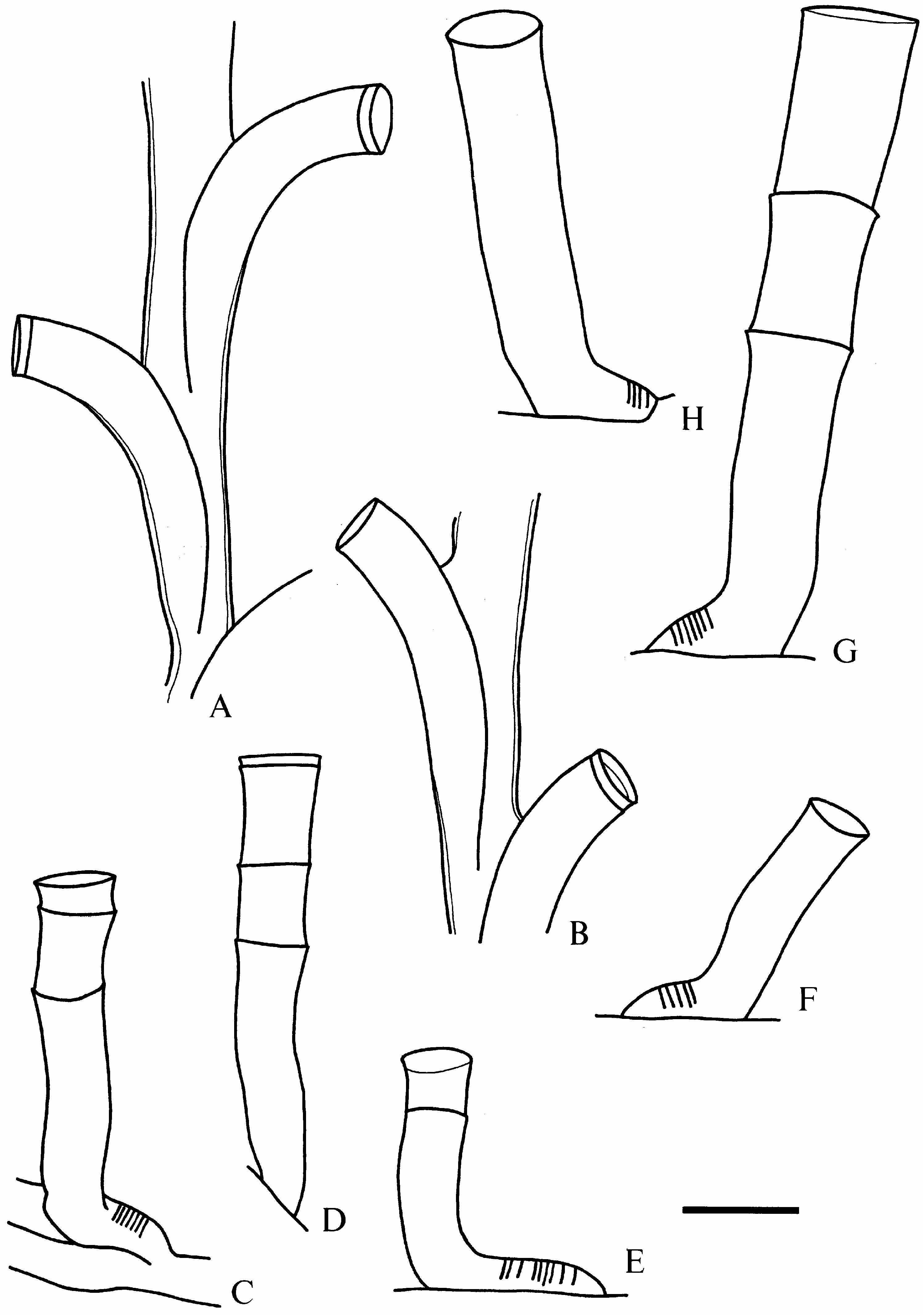
Campanularia antarctica Stechow, 1922 View in CoL
( figure 1I View FIG )
Campanularia volubilis var. antarctica Ritchie, 1913: 22–23 View in CoL , figure 6 View FIG .
Campanularia antarctica Stechow, 1922: 96 View in CoL ; 1923a: 104; 1923b: 3; Briggs, 1938: 17–18; Rees and Thursfield, 1965: 89, 195; Smaldon et al., 1976: 17.
? Campanularia volubilis var. antarctica: Blanco, 1965: 154 View in CoL , pl. 1 figures 2 View FIG , 6 View FIG , 7.
Material examined. ANT VIII- 5 Stn 16-459, several hydrothecae on Halecium delicatulum (RMNH-Coel. 30759).
Description. Stolonal hydrorhiza giving rise, by means of a short apophysis, to erect and slightly sinuous pedicels supporting a single hydrotheca resting on a basal spherule ca 40 M m high and 81 M m wide ( figure 1I View FIG ). Wall of pedicel smooth, but sometimes with a few distal annulations.
Hydrotheca bell-shaped ( figure 1I View FIG ), ca 725 M m high, with a diaphragm (ca 113 M m wide) delimiting a small basal chamber (ca 64 M m high). Distinct inflexion point present between basal chamber and remaining part of hydrotheca. Hydrothecal diameter strongly increasing after basal chamber; thereafter almost constant. Maximum diameter at aperture (ca 435 M m). Hydrothecal aperture circular; rim provided with ca 12 blunt cusps, separated by shallow embayments.
Gonothecae not found.
Remarks. Our material seems to be conspecific with the material described by Ritchie (1913) as Campanularia volubilis (Linnaeus) var. antarctica , though the absence of gonothecae prevents positive identification. It agrees with Ritchie’s material in almost every detail of trophosome structure. Thus, as in our material, Ritchie indicated that, in profile, the hydrothecal walls scarcely diverge from each other. The hydrothecal rim is provided with 10–13 rounded cusps. They also share the smooth perisarc of the hydrothecal pedicel, though in Ritchie’s material there are three to five twists at the base.
According to Totton (1930) there is a possibility that this species may prove to be identical with Hickson and Gravely’s (1907) material of Campanularia laevis Hartlaub, 1905 (now C. hicksoni Totton, 1930 ). Totton was inclined to regard Ritchie’s variety as a small Hickson and Gravely’s C. laevis . Rees and Thursfield (1965), however, considered C. hicksoni a much larger species than C. antarctica and proposed to keep them separate.
Blanco’s (1965) material has much smaller hydrothecae than those found by other authors and by ourselves. Moreover, though it has ca 12 cusps, these seem to be sharper. Furthermore, the basal chamber is longer. We have doubts whether or not this material belongs to C. antarctica .
Ecology and distribution. Campanularia antarctica has been found at depths from 13 ( Ritchie, 1913) to 108 m ( Briggs, 1938) on bottoms of small rocks with red algae ( Briggs, 1938); we found it at 350– 380 m. It has been reported epibiotic on Stegella lobata (cf. Ritchie, 1913) and we observed it on H. delicatulum . Ritchie (1913) found colonies with gonothecae in June and August.
Campanularia antarctica View in CoL seems to have a circum-Antarctic distribution. Hitherto, it was only known from East Antarctica: off Cape Royds, Ross Sea ( Ritchie, 1913) and at 65 ‡ 42’S, 92 ‡ 10’E ( Briggs, 1938). Our material, however, comes from the east coasts of the Weddell Sea (off Cape Norvegia, Princess Martha Coast).
Campanularia hicksoni Totton, 1930 View in CoL Campanularia hicksoni Totton, 1930: 148 View in CoL , figure 7a–e; Briggs, 1938: 15; Rees and
Thursfield, 1965: 90, 195; Blanco and Bellusci de Miralles, 1972: 10, figures 6 View FIG –19;
Naumov and Stepan’yants, 1972: 34, 37; Stepan’yants, 1972: 67, figure 12; 1979:
29, pl. 5 figure 1 View FIG ; Blanco, 1984: 18–20, pl. 13 figures 29–31; 1994a: 159; 1994b:
192.
? Campanularia hicksoni: El Beshbeeshy, 1991: 94–97 View in CoL , figure 22a.
Campanularia laevis: Hickson and Gravely, 1907: 25 View in CoL , pl. 4 figure 26; Vanhöffen, 1910: 298, figure 18; Ritchie, 1913: 19, figure 5 View FIG .
Material examined. ANT VII- 4 (Epos 3) Stn 274, several hydrothecae on Staurotheca sp. (MNHN-Hy 1285; RMNH-Coel. 30756); ANT VIII- 5 Stn 16- 454, several hydrothecae on Oswaldella sp. (RMNH-Coel. 30757); ANT VIII- 5 Stn 16-496, several hydrothecae on Oswaldella sp. (RMNH-Coel. 30758).
Remarks. In our material the length of the hydrothecae is 2335–2946 M m and the diameter at the aperture is 902–1417 M m. The length of the basal chamber is 161–242 M m and the diameter at diaphragm is 322–483 M m. The basal spherule is 97–113 M m high and 161–193 M m wide.
Totton (1930) considered that, besides the typical form of the species, there was a smaller form. According to Totton (1930: 148): ‘Hydrothecae of very variable size, deep, cylindrical; larger ones with longitudinal folds passing down from the emarginations of the folded and sinuous margin, which has up to 20 rounded teeth of the same shape as that of the depressions between. Smaller specimens with margin less folded; teeth much larger than intervening spaces with flatter tops and from 9 upwards in number’. The two forms differ in size of both hydrothecae and gonothecae. Totton left it for future studies to determine whether or not the two forms can be regarded as different species. To us it seems likely that Totton was dealing with two different species since there are large differences in the shape of the hydrothecal cusps and in the size of both hydrothecae and gonothecae.
Totton (1930) was also inclined to regard the material described by Ritchie (1913), as C. volubilis var. antarctica , as the male colonies of the small form. However, we consider that Ritchie’s material belongs to C. antarctica , a clearly distinguishable species, as it was already stated by other authors (e.g. Rees and Thursfield, 1965).
El Beshbeeshy (1991) assigned to C. hicksoni material with distinctly smaller hydrothecae (length 860–1160 M m and diameter at aperture 406–464 M m).
Ecology and distribution. Campanularia hicksoni has been found at depths from 10 ( Naumov and Stepan’yants, 1972) to 385 m ( Vanhöffen, 1910); we found it between 80 and 280 m. It has been reported on bottoms of shingle ( Totton, 1930), bottoms of small rocks with red algae ( Briggs, 1938), bottoms of stones, sponges, algae and hydroids; hydroids, sponges and algae; sand and stones bearing hydroids, sponges and bryozoans; gravel covered with hydroids and alcyonarians; muddy bottoms with bryozoans, hydroids and sponges; bottoms with abundant bryozoans and hydroids; muddy bottoms with numerous stones; bottoms with hydroids and sponges; rocky bottoms with Bryozoa and numerous stones, and stony and rocky bottoms ( Naumov and Stepan’yants, 1972). Campanularia hicksoni has mainly been found epibiotic on hydroids ( Hickson and Gravely, 1907; Ritchie, 1913; Totton, 1930; Briggs, 1938; Naumov and Stepan’yants, 1972; Stepan’yants, 1972), but also on caprellids ( Totton, 1930); we also found it on other species of hydroids ( Oswaldella sp. and Staurotheca sp. ). It has been found with gonothecae in January ( Totton, 1930) and February ( Hickson and Gravely, 1907).
Campanularia hicksoni View in CoL seems to have a circum-Antarctic distribution, being known from both East and West Antarctica. From the former, it has been reported from McMurdo Sound ( Hickson and Gravely, 1907; Ritchie, 1913), off Cape Royds ( Ritchie, 1913) and off Cape Adare ( Totton, 1930), all in the Ross Sea area, from the Davis Sea ( Vanhöffen, 1910; Briggs, 1938; Stepan’yants, 1972, 1979), from Commonwealth Bay ( Briggs, 1938) and d’Urville Sea ( Naumov and Stepan’yants, 1972), both off Adélie coast, and from the Friendship Sea ( Stepan’yants, 1979). In West Antarctica, C. hicksoni View in CoL was reported from Peter I Island ( Blanco and Bellusci de Miralles, 1972), Low Island ( Blanco, 1984) and the South Shetland Islands ( Stepan’yants, 1979). Our material comes from the east coast of the Weddell Sea (off Cape Norvegia and Atka Iceport, Princess Martha Coast).




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Filellum sp.
| Cantero, A. L. Peña, Svoboda, A. & Vervoort, W. 2004 |
Campanularia hicksoni: El Beshbeeshy, 1991: 94–97
| EL BESHBEESHY, M. 1991: 97 |
Campanularia volubilis var. antarctica: Blanco, 1965: 154
| BLANCO, O. M. 1965: 154 |
Hebella plana: Totton, 1930: 156
| BOERO, F. & BOUILLON, J. & KUBOTA, S. 1997: 13 |
| BLANCO, O. 1994: 161 |
| BLANCO, O. 1994: 190 |
| BLANCO, O. M. & ZAMPONI, M. O. & GENZANO, G. N. 1994: 21 |
| EL BESHBEESHY, M. 1991: 57 |
| BOERO, F. 1980: 134 |
| STEPAN'YANTS, S. D. 1979: 55 |
| SMALDON, G. & HEPPELL, D. & WATT, K. R. 1976: 15 |
| REES, W. J. & THURSFIELD, S. 1965: 74 |
| BRIGGS, E. A. 1938: 23 |
| TOTTON, A. K. 1930: 156 |
Lafoea dumosa: Totton, 1930: 158
| PENA CANTERO, A. L. & GARCIA CARRASCOSA, A. M. 1995: 23 |
| MILLARD, N. A. H. 1977: 15 |
| TOTTON, A. K. 1930: 158 |
Billardia subrufa: Totton, 1930: 151
| GENZANO, G. N. & ZAMPONI, M. O. 1997: 290 |
| BOUILLON, J. & MASSIN, C. & KRESEVIC, R. 1995: 78 |
| PENA CANTERO, A. L. & GARCIA CARRASCOSA, A. M. 1995: 131 |
| PENA CANTERO, A. L. 1991: 209 |
| STEPAN'YANTS, S. D. 1979: 39 |
| NAUMOV, D. V. & STEPAN'YANTS, S. D. 1972: 34 |
| STEPAN'YANTS, S. D. 1972: 67 |
| VERVOORT, W. 1972: 82 |
| BLANCO, O. M. 1967: 252 |
| NAUMOV, D. V. & STEPAN'YANTS, S. D. 1962: 71 |
| BROCH, H. 1948: 4 |
| BRIGGS, E. A. 1938: 18 |
| TOTTON, A. K. 1930: 151 |
Campanularia hicksoni
| BRIGGS, E. A. 1938: 15 |
| TOTTON, A. K. 1930: 148 |
Campanularia antarctica
| SMALDON, G. & HEPPELL, D. & WATT, K. R. 1976: 17 |
| REES, W. J. & THURSFIELD, S. 1965: 89 |
| BRIGGS, E. A. 1938: 17 |
| STECHOW, E. 1923: 104 |
| STECHOW, E. 1923: 3 |
| STECHOW, E. 1922: 96 |
Lytoscyphus subrufus:
| STECHOW, E. 1919: 82 |
Campanularia volubilis var. antarctica
| RITCHIE, J. 1913: 23 |
Lafoea gaussica Vanhöffen, 1910: 312
| STEPAN'YANTS, S. D. 1979: 48 |
| TOTTON, A. K. 1930: 159 |
| VANHOFFEN, E. 1910: 312 |
Hebella striata var. plana
| JADERHOLM, E. 1926: 4 |
| BILLARD, A. 1914: 9 |
| VANHOFFEN, E. 1910: 314 |
| RITCHIE, J. 1907: 530 |
Campanularia laevis:
| RITCHIE, J. 1913: 19 |
| VANHOFFEN, E. 1910: 298 |
| HICKSON, S. J. & GRAVELY, F. H. 1907: 25 |
Campanularia subrufa Jäderholm, 1904: 5
| NUTTING, C. C. 1915: 42 |
| VANHOFFEN, E. 1910: 296 |
| JADERHOLM, E. 1905: 15 |
| JADERHOLM, E. 1904: 5 |
Lafoea fruticosa: G. O. Sars, 1874: 114
| BLANCO, O. M. & ZAMPONI, M. O. & GENZANO, G. N. 1994: 27 |
| BLANCO, O. M. 1984: 15 |
| STEPAN'YANTS, S. D. 1979: 47 |
| VERVOORT, W. 1972: 66 |
| STEPAN'YANTS, S. D. 1972: 67 |
| NAUMOV, D. V. & STEPAN'YANTS, S. D. 1962: 76 |
| TOTTON, A. K. 1930: 157 |
| ALLMAN, G. J. 1888: 34 |
| SARS, G. O. 1874: 114 |
Lafoea gracillima: G. O. Sars, 1874: 115
| BRIGGS, E. A. 1938: 25 |
| TOTTON, A. K. 1930: 158 |
| STECHOW, E. 1925: 457 |
| BILLARD, A. 1914: 10 |
| VANHOFFEN, E. 1910: 312 |
| RITCHIE, J. 1907: 531 |
| JADERHOLM, E. 1905: 21 |
| HARTLAUB, C. 1905: 594 |
| HARTLAUB, C. 1904: 6 |
| JADERHOLM, E. 1903: 273 |
| SARS, G. O. 1874: 115 |
Lafoea capillaris G. O. Sars, 1874: 115
| SARS, G. O. 1874: 115 |
Campanularia gracillima
| ALDER, J. 1856: 361 |
Campanularia fruticosa M. Sars, 1851: 138
| SARS, M. 1851: 138 |
Sertularia dumosa
| FLEMING, J. 1820: 83 |